Nomenclature
Common sunflower (Helianthus annuus L.) belongs to the Asteraceae family (Schilling 2006).
Family
Asteraceae – Aster family
Genus
Helianthus
Species
annuus
NRCS Plant Code
HEAN3 (USDA NRCS 2026).
Subtaxa
There are no subtaxa recognized by the Flora of North America. Although the species is highly variable, taxonomists have yet to recognize intraspecific taxa (Schilling 2006).
Synonyms
Helianthus annuus subsp. jaegeri (Heiser) Heiser, H. a. subsp. lenticularis (Douglas ex Lindl.) Cockerell, H. a. subsp. texanus Heiser, H. a. var. lenticularis (Douglas ex Lindl.) Steyerm, H. a. var. macrocarpus (DC.) Cockerell, H. a. var. texanus, H. aridus Rydberg, H. jaegeri Heiser, H. lenticularis Douglas ex Lindley, H. macrocarpus DC (Schilling 2006).
Common Names
Common sunflower, annual sunflower, girasol, hairy-leafed sunflower, Kansas sunflower, mirasol (Schilling 2006; Wall and MacDonald 2009; Pavek et al. 2012; Spellenberg 2012; Kindscher 1987).
Chromosome Number
Chromosome number is: 2n = 34 (Schilling 2006).
Hybridization
Common sunflower hybridizes with other annual wild and crop Helianthus species (Schilling 2006; Lesica 2012). Hybridization with prairie sunflower (H. petiolaris) occurs and local hybrid swarms are possible in areas of species overlap (Cronquist 1994; Johnson and Larson 1999). Common sunflower also forms natural hybrids with silverleaf sunflower (H. argophyllus), serpentine sunflower (H. bolanderi), cucumberleaf sunflower (H. debilis), paradox sunflower (H. paradoxus), and Texas sunflower (H. praecox) (Heiser et al. 1969). Common sunflower × serpentine sunflower F1 hybrids studied in California were almost completely sterile (Carney et al. 2000).
Distribution
Common sunflower is native to western North America, but because of escaped crop, horticultural, and forage cultivars it now has a widespread North American and worldwide distribution (Schilling 2006; Hitchcock and Cronquist 2018). Hybridization and introgression have also contributed to the expanding distribution of common sunflower (Heiser et al. 1969). In the United States, it occurs in all states but Alaska. It is common on dry sites west of the Mississippi River, and grows outside of cultivation in eastern states (Harrington 1967; Munz and Keck 1973; Weber 1976; Schilling 2006; Hitchcock and Cronquist 2018).
Domestication and cultivation of common sunflower occurred prior to colonization (Lentz et al. 2001). Archaeological studies conducted between 1997 and 2000 recovered hulled common sunflower seeds aged to 4085 BP and unhulled seeds aged to 4130 BP at the San Andres site in the Gulf Coast region of Tabasco, Mexico (Lentz et al. 2001). There is also evidence of domestication dating back to 4625 BP in middle Tennessee (Crites 1993, cited in Seiler et al. 2017), which suggests very early domestication of common sunflower in the eastern US was not an independent occurrence (Lentz et al. 2001).
Habitat And Plant Associations
Widely distributed, common sunflower grows in a large variety of habitats and plant communities. Availability of open, disturbed sites is critical to species occurrence (Hickman 1993; Andersen and Holmgren 1996; Hazlett 1998; Schilling 2006; Welsh et al. 2015). It thrives in open, sunny, disturbed sites, and is often abundant in foothills, along roadsides and stream banks, and in old fields (Fig. 1) (Andersen and Holmgren 1996, Lesica 2012). On disturbed sites, common sunflower populations can range from a few to several thousand plants (Fig. 2) (Pilson 2000).

Figure 1. Common sunflower plant growing along a gravel roadside in sagebrush and pinyon-juniper woodland habitat in Colorado. Photo: USDI Bureau of Land Management (BLM) CO 130 Seeds of Success (SOS).

Figure 2. Large population of common sunflower along both sides of a roadway in New Mexico. Photo: USDI Bureau of Land Management (BLM) CA 018 Seeds of Success (SOS).
Plant associations discussed here represent just a fraction of the potential habitats where common sunflower grows. It made up to 17.8 lbs/ac (2 g/m2) of the standing biomass in northern mixed-grass prairie dominated by blue grama (Bouteloua gracilis), field brome (Bromus arvensis), and western wheatgrass (Pascopyrum smithii) (Karl et al. 1999). In Utah, common sunflower occurred in saltgrass-muhly (Distichlis–Muhlenbergia spp.) grassland, desert shrubland, pinyon-juniper (Pinus–Juniperus spp.) woodland, and mountain brush (Fig. 3) (Welsh et al. 2015). In San Juan County, Utah, common sunflower was a component of communities with greasewood (Sarcobatus vermiculatus), cheatgrass (Bromus tectorum), crested wheatgrass (Agropyron cristatum), Indian ricegrass (Achnatherum hymenoides), scarlet globemallow (Sphaeralcea coccinea), and prickly Russian thistle (Salsola tragus) on loamy fine sands at sites receiving 8 to12 in (200-300 mm) of annual precipitation (Tilley and Pickett 2021). In southern Nevada, common sunflower occurred in small widely scattered populations on disturbed sites in sagebrush (Artemisia spp.) and saltbush (Atriplex spp.) communities at 2,200 to 6,600 ft (670-2,000 m) elevations (Beatley 1976). In Canyon de Chelly National Monument, Arizona, common sunflower occurred in riparian areas but was classified as a mostly upland species (Reynolds and Cooper 2011). Common sunflower is noted as an important forb in blue grama-western wheatgrass and big bluestem-little bluestem (Andropogon geradii–Schizachyrium scoparium) rangeland types common in the northern High Plains in northeastern New Mexico and the Texas Panhandle (Shiflet 1994). On the Hawaiian island of Oahu, common sunflower occurred a distance of 130 to 160 ft (40-50 m) from the average sea water level along coastlines (Richmond and Mueller-Dombois 1972).

Figure 3. Large population of common sunflower growing in a woodland opening in California. Photo: USDI Bureau of Land Management (BLM) CA 180 Seeds of Success (SOS).
Elevation
Common sunflower occupies habitats ranging from 0 to 9,840 ft (3,000 m) in elevation throughout its range (Schilling 2006). This is also the elevation range for the species in the Sonoran Desert (Spellenberg 2012). Common sunflower is mostly found below 6,600 ft (2,000 m) in California (Hickman 1993; Keil 2020) and below 5,250 ft (1,600 m) in Oregon (Chambers 2020). In Utah, common sunflower grows at elevations from 3,940 to 8,005 ft (1,200-2,440 m) (Welsh et al. 2015).
Soils
Common sunflower grows in a variety of soil types but seems to prefer clay soils (Heiser 1947; Cronquist 1994). It occupies dry to moist soils and tolerates some metals and salt making it useful in mine reclamation (Cronquist 1994). It has been grown successfully in serpentine soils collected from the western United States (Walker 1954). In Saskatchewan grasslands, it was reported on unconsolidated sand dunes with low fertility (Looman 1963). It also occurred in saltgrass communities with 0.1% salinity soils near Lincoln, Nebraska (Ungar et al. 1969).
On restoration websites for Arizona and Utah, common sunflower is recommended for revegetation on clay, loam, silt, gravel, and sand soils and is considered to be tolerant of acidic and alkaline conditions, metals, and salt (Fig. 4) (University of Arizona 2026, Utah State University 2025).

Figure 4. Small stature common sunflower growing in gravel in Oregon. Photo: USDI Bureau of Land Management (BLM) OR 010 Seeds of Success (SOS).
Description
Common sunflower is a highly variable annual (Figs. 4 and 5), considered the most variable of the Helianthus species with size and form being dependent on environment and distribution (Andersen and Holmgren 1996). Plants are stout, erect, usually branched, and range from 3 to 10 ft (1-3 m) tall (Johnson and Larson 1999; Spellenberg 2012; Welsh et al. 2015; Chambers 2020; Keil 2020; LBJWC 2025). They are taprooted with numerous fibrous roots (Currah et al. 1983). A maximum rooting depth of 8.9 ft (2.7 m) was reported in silt loam soils in Nebraska (Weaver 1926 cited in Canadell et al. 1996). Herbage is usually sparsely covered with sharp stiff hairs (Spellenberg 2012) creating a gray green pubescence with a sandy, papery texture (Andersen and Holmgren 1996).

Figure 5. Large stature common sunflower growing on the Colorado Plateau. Photo: USDI Bureau of Land Management (BLM) CP 3 Seeds of Success (SOS).
Plants may have a single stem but more commonly have multiple branches arising from the upper half of the main stem. Stems terminate with a single large showy flower head (Pavek et al. 2012). Leaves low on the main stem are opposite, but the majority of the leaves occur on the upper part of the plant and alternate (Pavek et al. 2012). Petioles are 0.8- to 8-in (2-20 cm) long. Blades are somewhat triangular and basally cordate; margins are toothed and may be gland dotted (Fig. 6). Leaf blades measure 2 to 16 in (5-40 cm) long and 0.8 to 16 in (2-40 cm) wide (Pavek et al. 2012; Welsh et al. 2015; Chambers 2020).

Figure 6. Close-up of common sunflower’s hairy stems and somewhat cordate leaves. Photo: USDI Bureau of Land Management (BLM) WY 080 Seeds of Success (SOS).
Terminal flower heads are large, 3 to 5 in (8-13 cm) wide, and typically numerous (Pavek et al. 2012). At the University of Nebraska’s Northeast Cedar Point Biological Station where common sunflower is abundant, plants produced 10 to 18 inflorescences depending on crowding and water availability (Pilson 2000). Flower heads are comprised of many sterile yellow ray florets (17-30+) measuring up to 2 in (5 cm) long that surround 150 or more reddish to dark brown disk florets, which are perfect and fertile (Figs. 7 and 8) (Cronquist 1994; Pavek et al. 2012). Common sunflower fruits are cypselas that are often, yet incorrectly, referred to as achenes (LBJWC 2025). Seeds are 3 to 15 mm long with pappus scales that are 2 to 3.5 mm long (Chambers 2020). Under a hand lens, seeds are 4-angled (Craighead et al. 1963). Seeds are generally grayish with dark lengthwise streaks and spotting (Fig. 13) (Martin and Barkley 2000).

Figure 7. Common sunflower with about 20 yellow ray flowers surrounding dark brown disk flowers. Photo: USDI Bureau of Land Management (BLM) CA 320 Seeds of Success (SOS).

Figure 8. Common sunflower with less than 20 yellow ray flowers surrounding reddish-brown disk flowers. Photo: USDI Bureau of Land Management (BLM) CA 320 Seeds of Success (SOS).
Distinguishing wild common sunflower. Throughout this species review, unless otherwise noted, common sunflower refers to the wild plant. Wild plants refer to the native species that has not hybridized with crop plants, which are grown commercially for oilseed, food, and as ornamentals. Crop-wild hybrids are those plants where mixing of wild and crop plants has occurred or is occurring. Many studies describe the morphological and phenological differences between wild, crop, and crop-wild common sunflower populations. In a review by Dechaine et al. (2009), when mixing occurs, traits of crop plants can persist in wild populations for five generations or more. Wild plants are shorter with narrower leaves and stems, many more branches, smaller and more numerous flower heads, and they flower later and longer than crop or crop-wild hybrid plants (Table 1) (Reagon and Snow 2006; Dechaine et al. 2009).
Table 1. Traits of wild, crop-wild hybrids, and crop common sunflower plants grown from distinct populations collected in Colorado and grown in a common garden in Columbus, Ohio (Reagon and Snow 2006).
| Plant type | No. of branches | No. of flower heads | Diameter of first flower head |
| Wild | 22.7a | 267.5a | 46.0a |
| Crop-wild | 16.3b | 136.8b | 116.3b |
| Crop | 6.0c | 20.1c | 191.2c |
In a greenhouse study, wild common sunflower plants grew significantly more slowly (P < 0.01) and were less affected by drought, herbivory, and fungal infection treatments than crop plants (Mayrose et al. 2011). Wild accessions came from seed collected from native populations in Saskatchewan, Utah, Colorado, New Mexico, Arkansas, Kansas, and Iowa. Seed was germinated in a laboratory and then grown in a greenhouse (72-77 °F [22-25 °C] and 16 h/8 h light/dark cycle). Treatments were applied about 4 weeks after germination when seedlings had 4 to 8 true leaves (Mayrose et al. 2011).
Belowground Relationships And Interactions
Common sunflower roots in disturbed areas of Utah were colonized by vesicular-arbuscular mycorrhizae (Pendleton and Smith 1983). In a greenhouse experiment, common sunflower roots were colonized by mycorrhizae but the relationship did not affect plant growth (Wilson and Hartnett 1998).
Reproduction
Common sunflower reproduces exclusively from seed. Flowering can occur as early as February and as late as October depending on site and habitat conditions (Munz and Keck 1973; Spellenberg 2012; Keil 2020). Flowering from March to October is reported in the Sonoran Desert (Spellenberg 2012).
Breeding System
Common sunflower is primarily self-incompatible; cross pollination is generally required to produce viable seed (Heiser et al. 1969). Ellstrand et al. (1978) found that outcrossing indices varied from 0.54 to 0.91 (x = 0.76) in six large wild common sunflower populations in eastern Kansas. Outcrossing indices were inversely correlated with plant density so as plant density increased the rate of outcrossing decreased.
Pollination
There are hundreds of species of pollinators associated with common sunflower (Discover Life 2024). For more on pollinator use see Wildlife and Livestock Use.
Ecology
Common sunflower is a ruderal species typically most abundant on open, disturbed sites (Goodrich and Neese 1986; Johnson and Larson 1999; Pavek et al. 2012; Keil 2020). It is an early colonizer of roadsides, ditch banks, deteriorated rangelands, abandoned crop land, and other waste areas (Johnson and Larson 1999; Pavek et al. 2012). Spread by seed is rapid, especially on disturbances where competition is limited (LBJWC 2025).
Seed And Seedling Ecology
Common sunflower is a prolific seed producer (Fig. 9) (Stevens 1932) that provides a seed source for many insects and small mammals (Pilson 2000). It produces a seed bank and reproduces best at disturbed sites (Teo-Sherrell 1996).
Seed production. A single, well-developed plant growing with minimal competition in North Dakota evaluated at the time of likely maximum seed production had 7,200 seeds (Stevens 1932). In a census of roadside populations (n=21) of common sunflower near Lawrence, Kansas, the density of flower heads averaged 1.5/ft2 (16.6/m2) and the number of seeds/head averaged 107 based on counts from 24 individual seedheads at one site (Cummings and Alexander 2002).

Figure 9. Common sunflower plants with numerous seedheads in California. Photo: USDI Bureau of Land Management (BLM) CA 320 Seeds of Success (SOS).
Single plants growing in Nebraska produced between 1,400 and 16,100 seeds based on year and plant densities in a controlled field experiment (Teo-Sherrell 1996). Seed collected from wild plants was grown in plots with various plant densities (1993): plants without conspecific neighbors or in patch densities of 0.02/ft2 (0.25/m2) or 0.9/ft2 (1/m2). In the next growing season (1994), all plants were grown in 0.02/ft2 (0.25/m2) patches. Total seed produced/plant ranged from 5,700 to 14,500 in 1993 and from 1,400 to 16,100 in 1994. Average seed production/plant was 5,800 for plants lacking conspecific neighbors, 12,300 for plants with the nearest neighbor 16 ft (5 m) away, and 8,200 for plants in patches. More than 85% of seed was filled, with less than 10% empty and less than 6% damaged. Weevils and lepidopteran larvae were observed, but bird predation was rare. At the time when seed counts were made, seed was not yet dehydrating and remained firmly attached in the head, which may have made removal difficult for birds (Teo-Sherrell 1996).
In an experimental field study, seeding density had little impact on seed production of common sunflower (Cummings and Alexander 2002). Plots (6.6 ft [2m] × 6.6 ft [2 m]) seeded with a high density of seed (10,000 seeds/plot) produced significantly more seedlings and adult plants than those seeded at a low density (2,000 seeds/plot), but seed production estimates were not significantly different between plots. Treatments designed to exclude birds and small mammals resulted in more stems in protected plots but did not significantly affect seed production (Cummings and Alexander 2002).
Seed predation. Flowers and seeds of common sunflower are subject to predation by insects, birds, and mammals. At the University of Nebraska’s Cedar Point Biological Station, the most abundant and damaging insect predators were the sunflower bud moth (Suleima helianthana), sunflower moth (Homoeosoma electellum), head-clipping weevils (Haplorhynchites aeneus), and seed weevils (Smicronyx fulvus and S. sordidus) (Pilson 2000).
Analyses of seed predation impacts often include impacts or levels of damage from seed predation have often included wild and crop-wild plants. Cummings et al. (1999) found that crop-wild hybrids received more insect damage than wild plants in a field experiment at the University of Kansas’ Ecological Reserves. An average of 36.5% of crop-wild plant’s seeds were eaten by insect larvae, compared to 1.8% of wild plant’s seeds. Differential feeding by insects could have been related to phenology or plant traits. Crop-wild plants flowered earlier (average 20 days) and produced fewer flower heads and seeds than wild plants. Few of the wild plants were flowering at the time of peak insect damage. Crop-wild plants received more damage from weevils and sunflower moths, and wild plants had more damage from banded sunflower moths (Cummings et al. 1999).
Using the same crop-wild and wild plants as above, researchers looked at predation by small mammals (Alexander et al. 2001). Seeds produced by crop-wild plants were about twice as big as seed produced by wild plants. Seed was set out in the field on 5 March, 23 April, and 19 May for 10 days. More crop-wild (62.3%) than wild (41.7%) seed was taken. Dominant seed predators in the region were western harvest mice (Reithrodontomys megalotis) with fewer numbers of white-footed mouse (Peromyscus leucopus) and prairie voles (Microtus ochrogaster) (Alexander et al. 2001).
Predation was higher for wild than crop-wild and crop common sunflower seed early in feeding trials in sideoats grama (Bouteloua curtipendula)-little bluestem grassland at the University of Nebraska’s Cedar Point Biological Station (Dechaine et al. 2010). Rodent predation was predominant. In the first 2 days of setting out seed: 11.3% of wild, 6.2% of crop-wild, and 1.3% of crop seed was removed. By day 6, predation was similar across seed types (38-47%). Predation on day 4 and day 6 was positively correlated with the oil and energy content of seeds, which was greater for crop seeds. Researchers suggested granivores may prefer seed they recognize (wild) initially but will preferentially remove unknown seed with high oil or energy content (Dechaine et al. 2010).
Seed banking. Common sunflower produces a seed bank, and studies indicate some seed remains viable after 4 or more years of burial (Alexander and Schrag 2003) but this is likely a small proportion of the year’s seed (Toole and Brown 1946). Toole and Brown (1946) reported on early and later findings of seeds buried by J.W.T. Duvel. Seeds were buried at various depths beneath heavy clay soils at Arlington Experimental Farm in Alexandria, Virginia. Seed was put in pots and buried on December 19, 1902. Pots were dug in November 1903. Seed from pots buried 8 in (20 cm) deep germinated at 44%, 22 in (56 cm) deep at 64% and 42 in (107 cm) deep at 67%. No germination came from pots buried longer than a year (Toole and Brown 1946).
Teo-Sherrell (1996) found that 78% of common sunflower seeds were still viable after 1 year of burial. Experiments were conducted in Lincoln, Nebraska, where winters are cold and dry, springs are mild and moist, and summers are hot and humid. Fresh seed was collected and buried in soil in November 1992 and 1993 at depths of 1 to 5 in (2.5-12.7 cm) or 5 to 9 in (12.7 to 22.9 cm). Depth of burial did not affect survival (Teo-Sherrell 1996).
Alexander and Schrag (2003) used field studies to evaluate seed banking in common sunflower populations. Seed used in the study was wild collected from plants growing in Lawrence, Kansas. Fiberglass mesh seed packets with 140 seeds each were buried 10 in (25 cm) deep or left on the soil surface at two sites at the Kansas Field Station and Ecological Reserves in Jefferson County (Alexander and Schrag 2003). On average, 47% of seed was viable 4 years after fresh seed was buried in November 1999 and 16% was viable after 3 years when fresh seed was buried in December 2000. For seed not buried at left on the soil surface, 11 to almost 30% produced seedlings in the first year, and seedlings were rare in year two. In a companion field study, seed inputs were manipulated by only allowing a subset of plants to produce seed and removing immature heads as they appeared on other experimental plants and those up to 13 ft (4 m) from the experimental site. Plants allowed to disperse seed produced and average of 176 seeds/head and 22,964 seeds/plant, and researchers estimated that about 23% of seedlings originated from soil-stored seed (Alexander and Schrag 2003).
Seed banks were important to population dynamics of rural roadside common sunflower populations in eastern Kansas (mean annual ppt: 39 in [1000 mm]) and western Nebraska (mean annual ppt: 17 in [432 mm]) (Alexander et al. 2009). Plant densities were generally higher in Nebraska than in Kansas and abundance was linearly correlated with spring precipitation in Nebraska but not in Kansas. Stretches of roads with 100s to 1000s of plants often had virtually no plants present the year prior but large numbers of plants 2 years prior. Colonization occurred near sites where populations existed the previous year, for 2 years in Nebraska and for 1 year in Kansas, but more than 25% of colonization appeared at least 525 ft (160 m) from the nearest possible seed source population much farther than the typical seed dispersal distance of 3 ft (0.9 m) (Alexander et al. 2009).
Seedling emergence from the seed bank increases with disturbance and increases even more when soil is disturbed before and after seed dispersal (Moody-Weis and Alexander 2007). Researchers determined this through a field experiment conducted at Nelson Environmental Studies Area in northeastern Kansas (mean annual ppt: 37 in [930 mm]). The field site was a grassland on clay loam soils dominated by smooth brome (Bromus inermis), tall fescue (Schedonorus arundinaceus), and Kentucky bluegrass (Poa pratensis). Seedbank plots (50 × 50 in [127 × 127 cm]) were seeded at three densities: no seed added (control), 1,700 seeds added (low density), and 2,600 seeds added (high density) and subjected to three levels of soil disturbance: none (control), small (vegetation removed and top 4 in (10 cm) of soil turned over in patches leaving about 9% undisturbed), large (all vegetation removed and top 4 in (10 cm) of soil turned over throughout), and disturbed at various times: before (2 wks before seeding), after (1-2 days after seeding), or both before and after seeding. Seed was added and disturbances applied just once in 2001. In 2003 and 2004, all plots were tilled prior to seedling emergence to estimate seed banks. Soil disturbances significantly increased emergence regardless of size or timing, but soil disturbances that occurred before and after seed dispersal had greater emergence than those disturbed once. Disturbances that occurred after seed dispersal increased the likelihood of reproduction and resulted in large seed bank formation which remained after 2 years of germination. Seed density did not affect seed bank formation but did result in more emergence in plots that were disturbed after seeding (Moody-Weis and Alexander 2007).
Disturbance Ecology
Common sunflower is an early colonizer of disturbed and open sites. Occurrence and abundance are often associated with degree of disturbance or harsh conditions (Tolstead 1941b; Albertson and Weaver 1944; Weaver 1968). In prairie regions, common sunflower is considered important to soil stabilization of denuded soils or sites made bare by drought or wind erosion (Weaver 1968). When Midwest prairies experienced 8 years of severe drought, common sunflower abundance increased as dominant prairie species decreased. In drought times, common sunflower covered large areas of degraded range lands, although plants rarely exceeded 4 ft (122 cm) tall (Albertson and Weaver 1944).
Old field succession. Common sunflower is a usual volunteer soon after the abandonment of agricultural land and often persists several years into secondary succession (Tolstead 1941b). When abandoned cropland and relatively undisturbed grasslands were compared in southeastern Washington, common sunflower occurred in 1- to 52-year-old abandoned fields but not in undisturbed grasslands dominated by bluebunch wheatgrass (Pseudoroegneria spicata) and Sandberg bluegrass (Poa secunda). Cover of common sunflower was 3% in 2-year-old, 29% in 5-year-old, and 8% in 12-year-old fields where rubber rabbitbrush (Ericameria nauseosa) and cheatgrass were common associates (Daubenmire 1975).
In the first year of agricultural abandonment in the Black Forest of Colorado’s Piedmont, common sunflower dominated the annual weed stage, and it persisted but with lower abundance in fields abandoned for 4 years (Livingston 1949). In a survey of 70,000 acres (280 km2) of abandoned agriculture land in northern Weld County, Colorado, common sunflower was often the first plant to appear in locations not inundated by early spring flooding. Common sunflower frequency averaged 24% in fields abandoned for 5 years and decreased to 4% at 30 years after abandonment when fields were transitioning to native short-grass prairie (Costello 1944).
Fire and other overstory removal. Common sunflower occurs on burned sites soon after fire throughout its western range (Wright and Bailey 1980).
In the Black Hills of South Dakota, common sunflower biomass productivity was 131 lbs/ac (147 kg/ha) in the second growing season after clearcutting and burning in a ponderosa pine stand on a limestone shelf. Common sunflower seed was not recovered in seed bank sampling conducted before treatments (Wienk et al. 2004).
Fautin (1946) reported that common sunflower invades and occupies sites soon after sagebrush is burned or removed by other means in western Utah. It is also typical of early post-fire succession in pinyon-juniper communities in Utah (Pieper and Wittie 1990). In west-central Utah, common sunflower was an abundant forb in early post-fire succession, reaching maximum cover of 3.3% and frequency of 20% in the third post-fire year. Common sunflower was not present in older burns. This study evaluated 28 burns (3-100+ years old) in 17 locations of western juniper-singleleaf pinyon (Juniperus occidentalis–Pinus monophyla) vegetation in west-central Utah (Barney 1972). Common sunflower occurred in 1-year-old burned sites but not in 2-, 3-, or 4-year-old burned sites in Gambel oak (Quercus gambelii) vegetation near Salt Lake City, Utah (McKell 1950). Similarly, common sunflower was present on 1-year old burned big sagebrush (Artemisia tridentata) and Utah juniper-singleleaf pinyon (J. osteosperma–P. edulis) sites in west-central Utah but not on 2- or 3-year-old burns or adjacent unburned sites (Ott et al. 2001).
In Colorado, common sunflower established the first growing season after fire in arid salt desert BLM lands within the McInnis Canyons National Conservation Area and Grand Junction Field Office (personal comms cited in Grant-Hoffman et al. 2003). It was also predominant in early post-fire succession (within 2 years of fire) in Utah juniper-singleleaf pinyon habitats on highly eroded plateaus of southwestern Colorado’s Mesa Verde (Erdman 1970).
On the Apache-Sitgreaves National Forest in east-central Arizona, common sunflower did not occur in ponderosa pine (P. ponderosa) forests before burning or thinning and burning treatments, but it did occur at a low frequency 5 years following treatments (Strahan et al. 2015). In singleleaf pinyon and juniper woodlands from western to east-central Arizona, common sunflower production increased from a trace to 14 lbs/ac (16 kg/ha) after the overstory was killed by girdling and sawing. Production represents an average of measurements taken 4, 6, and 8 years after treatment (Clary and Jameson 1981). Findings were somewhat different on the Fort Stanton Experimental Range in southern Lincoln County, New Mexico. Common sunflower basal cover was greater (1.2%) on untreated than on treated (0.5%) sites visited 10 or more years after pinyon and juniper trees were mechanically controlled by dragging a cable between tractors. Cover differences were not significant (Rippel et al. 1983).
Wildlife And Livestock Use
Common sunflower is an important wildlife species. It is highly palatable early in the growing season, and although palatability decreases with maturity, large mammals and livestock readily consume its flower heads (Hermann 1966; Johnson and Larson 1999). Common sunflower seeds are sought out and eaten by a variety of small mammals and birds (Michael and Beckwith 1955; Flinders and Hansen 1972; Johnson and Larson 1999; Welsh et al. 2015).
Ungulates. In the western United States, common sunflower is eaten by elk (Cervus elaphus), deer (Odocoileus spp.), and pronghorn (Antilocapra americana). Elk fed on common sunflower leaves in Yellowstone in July (Skinner 1928). Common sunflower made up 13% of the relative density of elk and 48% of mule deer (O. hemionus) diets based on feces collections in northern Utah range from January through March 1983 (Mower and Smith 1989). In ponderosa pine and twoneedle pinyon-oneseed juniper (J. monosperma) woodlands at the Los Alamos National Laboratory in north-central New Mexico, common sunflower was up to 3% of fall elk diets and up to 7% of summer mule deer diets although it was a very rare species in the study area (Sandoval et al. 2005). In north-central New Mexico’s San Antonio Mountains, common sunflower cover was just 1% in the winterfat-blue grama (Krascheninnikovia lanata–Bouteloua gracilis), but it made up 5% of deer, 3% of elk, 13% of pronghorn, and 11% of horse diets (Stephenson et al. 1985a). In this region, use of common sunflower by pronghorn increased in a drought year. Spring and summer diets included significantly more common sunflower (P <0.05) in 1980 (an extreme drought year) than in 1981 (a normal precipitation year) (Stephenson et al. 1985b).
Small mammals. Common sunflower seeds are eaten by many small mammals and are often used as a food source in small mammal trapping studies (Ransome et al. 2004; Sullivan et al. 2017; Bani-Yaghoub and Reed 2018). In shortgrass prairie in northeastern Colorado, common sunflower made up to 5% of the dry weight of plants in summer diets of white-tailed jackrabbits (Lepus californicus) and up to 3% of black-tailed jackrabbits (L. townsendii) summer diets (Flinders and Hansen 1972). In confined feeding trials of hispid cotton rats (Sigmodon hispidus), common sunflower was the second most preferred food (Michael and Beckwith 1955). In mesquite (Prosopis spp.) plains of Fisher County, Texas, common sunflower seeds were an important winter food of hispid cotton mouse (Chaetodipus hispidus), making up 10.3% of the volume of winter diets. It also made up 5.2% of the volume of winter and 3.7% in spring diets of Ord’s kangaroo rat (Dipodomys ordii) (Alcoze and Zimmerman 1973).
Birds. Many bird species feed on common sunflower seeds, and the descriptions of utilization below are likely just a sample of the types and degree of use. In Okanogan County, Washington, common sunflower seed was found in in 43% of crops and made up 4% of the crop volume of California quail (Lophortyx californicus) collected in the fall (Knight et al. 1979). It was found in 14% of fall-collected and 8.3% of winter-collected crops but made up a very small volume of ring-necked pheasants (Phasianus colchicus) diets. Common sunflower seed was found in 14.3% of fall-collected crops but made up a very minor volume of chukar partridge (Alectoris chukar) diets (Knight et al. 1979).
Based on observations made from June 22 to July 25 in Willow Creek Valley, Malheur County, Oregon, Brewer’s sparrow (Spizella breweri) often fed on common sunflower (Peck 1911). In central Montana, common sunflower frequency was 1 to 2% in summer greater sage-grouse (Centrocercus urophasianus) brood habitats (Peterson 1969). In agricultural areas of Utah County, Utah, common sunflower made up a large portion of seeds recovered from stomachs of 3 of 45 harvested ring-necked pheasants (Cottam 1929). Common sunflower grew in recently cultivated sites within a short-grass natural area in Aurora, Colorado, where horned larks (Eremophila alpestris) preferentially fed (Kelso 1931).
Common sunflower was considered an important food of wild turkeys (Meleagris gallopavo) on the San Carlos Indian Reservation in southeastern Arizona. This was based on analysis of crops of 23 turkeys killed between October 16 and November 15 and feces collected in December and January (Murie 1946). Common sunflower made up 0.8% volume and 1.0% mass of digestive tract contents and was present in 12.5% of the digestive tracts of female cinnamon teals (Anas cyanoptera) taken from the Bosque del Apache National Wildlife Refuge in New Mexico. Cinnamon teals were collected during spring migration (Thorn and Zwank 1993). Although common sunflower was uncommon in a study area of the southern Great Plains in Lea County, New Mexico, it was the 18th most common by volume and 14th most common by frequency in fall and winter food in diets of scaled quail (Callipepla squamata) based on crop analysis of 227 birds (Campbell et al. 1973).
In confined feeding trials for northern bobwhites (Colinus virginianus) conducted at the Southern Plains Range Research Station in Woodward, Oklahoma, common sunflower was the third most preferred food (Thacker and Springer 2016). Based on crops from 30 birds taken from west Texas in 1971, 1972, and 1973, common sunflower seeds were 4.8% by frequency, 3.3% by weight, and 2.8% by volume of the diets of lesser prairie chickens (Tympanuchus pallidicinctus) (Crawford and Bolen 1976).
Insects, Pollinators. The worldwide Discover Life database indicates that there are 55 distinct families and hundreds of species of pollinators, insects, fungi, and pathogens associated with common sunflower. This includes at least four families and more than 28 bee species (Discover Life 2024). Common sunflower is a host to the following butterflies and moths in North America: Aethes straminoides, Agrotis ipsilon, Amorbia effoetana, Autofrapha gamma, Chlosyne lacinia, Charidryas gorgone, Choristonerura rosaceana, Cochylis arthuri, C. hospes, Ctenoplusia oxygramma, Euxoa ochrogaster, Grammia arge, Helicoverpa sea, Heliothis virescens, Hoeosoma electellum, Manduca muscosa, Ogdoconta cinereola, Ostrinia nubilalis, Pelochrista rorana, P. womonana, Peridroma saucia, Phtheochroa lavana, Plagiomimicus spumosum, Pseudoplusia includens, P. unipuncta, Rachiplusia ou, Scrobipalpalu artemisiella, Spilosoma virginica, Spodoptera exigua, S. eridania, S. frugiperda, Suleima baracana, S. helianthana, S. lagopana, and Vanessa cardui (Graves and Shapiro 2003; Robinson et al. 2024).
Nutritional Value
Common sunflower seeds provide nutrition and energy for wildlife and humans. Johnson and Robel 1968) reported fat content of seeds at 26% of dry weight; SER, INSR, SID Kew (2026) reported seed oil content as 42%. Plants collected from northern bobwhite habitats in south Texas contained 16% crude protein, 36% crude fiber, 3.4% ash, 0.5% calcium, and 0.35% phosphorus (Wood et al. 1986).
The energy content of common sunflower seeds collected in early October 1966 from northeastern Kansas averaged 5,573 cal/g and ranged from 5,575 to 5,619 cal/g based on the collection site, which included lowlands, uplands, limestone breaks, and clay uplands (Johnson and Robel 1968). In a study by Simms (1985) energy content of common sunflower seeds averaged 3,650 Cal/kg. Seeds were collected from locations in Nevada and Utah. Seeds were 16% protein, 70% carbohydrates, 3% fat, 3% ash, and 9% moisture.
Common sunflower crops can be used as livestock silage and improved varieties have 90 to 95% the feed value of corn silage with higher protein and considerably more fat (KSU n.d.).
Ethnobotany
Common sunflower was and continues to be used by Indigenous people as a food staple and medicinal and ceremonial plant. Uses described here do not represent a full account of this widely utilized plant and crop. Several review documents provide good summaries of Indigenous uses and farming of common sunflower (Craighead, Craighead et al. 1963; Shay 1986; Moerman 2003; Kindscher 1987).
Food. The seeds, stems, flowers, and tubers of common sunflower are edible, and many Indigenous people farmed the plant, which they also used to feed their livestock (Gilmore 1919; Vestal 1952; Hart 1976; MacKinnon et al. 2014; LBJWC 2025). Seeds were beaten out of the flower heads into baskets (Chamberlin 1911; Castetter and Opler 1936). Seeds were processed by cracking the seed coats then soaking them in water where hulls floated to the top and seeds dropped to the bottom (Harrington 1967). Seeds were eaten raw or dried or roasted and ground into a meal mixed with bone marrow or grease for cakes or into a flour for breads. Crushed seeds boiled in water released oil which was skimmed off the top and used in cooking (Gilmore 1919; Castetter and Opler 1936; Johnson and Larson 1999; MacKinnon et al. 2014). Roasted and ground seed was also used to make a drink like coffee (Harrington 1967). Indians of the Missouri River region of North Dakota traveled with sunflower cakes made from ground seed mixed with grease (Hart 1976). Common sunflower stems produce a creamy white juice that the Paiute in Utah chewed like gum (Palmer 1878). The Arikara, Hidatsa, and Mandan tribes used cultivated common sunflower seeds in trading (Gilmore 1919).
Medicine. Common sunflower has many medicinal uses. Flowers and oils were used to treat coughs and other pulmonary troubles. Flowers as a poultice were used to treat burns (MacKinnon et al. 2014; LBJWC 2025). Crushed root or leaves made into a poultice were used to relieve blisters or chewed and applied to sores, swellings, and rattlesnake or spider bites after sucking the venom (Turner et al. 1990; MacKinnon et al. 2014; LBJWC 2025). Tea from leaves was taken for high fevers. Seeds and leaves were used as a diuretic and expectorant (MacKinnon et al. 2014; LBJWC 20250. First Nations people of Canada used common sunflower to treat rheumatism, headaches, kidney weakness, malaria, worms, fatigue, swellings, and blisters (MacKinnon et al. 2014).
Planting common sunflower near tribal homes was believed to provide protection from malaria (LBJWC 2025). Oils from the seeds were used as a hair and skin treatment (Johnson and Larson 1999). Ramah Navajo burned common sunflower pith on scratched warts to remove them (Vestal 1952).
The plant has hypoglycemic activity (MacKinnon et al. 2014). Common sunflower was used by pharmacists in Europe as early as 1568 for many of the aforementioned ailments (Hart 1976).
Additional uses. There were many other Indigenous uses of common sunflower reported in the literature. Flowers and seeds were used to make dyes (Craighead et al. 1963; Spellenberg 2001; LBJWC 2025). The Navajo used common sunflower in a liniment during the War Dance. The Navajo also used hollow stalks as prayer sticks and flutes in the Mountain Chant. Plants were also used in conjunction with horsehair traps to capture birds (Elmore 1944; Vestal 1952).
Horticulture
Common sunflower has been cultivated as a crop and horticulture plant worldwide for centuries. It is grown for seed, oil, livestock feed, and as an ornamental plant. Seed is widely available commercially for a very large variety of cultivars (Heiser et al. 1969; Andersen and Holmgren 1996; Dorn and Dorn 2007; LBJWC 2025) with multiple color variations and with plant and seed head size ranging from dwarf to giant (Dorn and Dorn 2007). They are hardy to USDA Zone 3, easy to grow, drought tolerant, and attractive to pollinators (Dorn and Dorn 2007; Oregon State University 2026).
Revegetation Use
Many growth characteristics and tolerances of common sunflower make it a useful revegetation and reclamation species. It grows in all soil textures and tolerates acidic and alkaline conditions, metals, and salts (University of Arizona 2026; Utah State University 2026). It establishes and reproduces quickly and provides erosion control, pollinator and wildlife habitat, and forage (Jorgensen and Stevens 2004; Ogle et al. 2013, 2019; University of Arizona 2026; Utah State University 2026).
Common sunflower is often recommended for pollinator habitat and other revegetation projects at sites receiving 6 to 16 in (150-410 mm) average annual precipitation (Ogle et al. 2013, 2019; Tilley et al. 2019). Near Twin Falls, Idaho, bees were found on common sunflower growing just outside of a burn scar suggesting it provides blooms for ground nesting bees that survive fire in sagebrush-juniper woodlands (Cane 2012).
Developing A Seed Supply
For restoration to be successful, the right seed needs to be planted in the right place at the right time. Coordinated planning and cooperation is required among all partners to select appropriate species and seed sources; these are determined by site conditions and current or projected revegetation requirements and goals (PCA 2015) and consultation with land managers and stakeholders.
Developing a seed supply begins with seed collection from native stands. Collected seed is distributed for restoration or production of nursery stock. Increase in agricultural seed fields may be required to provide a supply of seed (PCA 2015). Production of nursery stock requires less seed than large-scale seeding operations. Regardless of the size and complexity of any revegetation effort, seed certification is essential for tracking seed origin from collection through use (UCIA 2015).
Seed Sourcing
Because empirical seed zones are not currently available for common sunflower, generalized provisional seed zones developed by Bower et al. (2014) may be used to select and deploy seed sources. These provisional seed zones identify areas of climatic similarity with comparable winter minimum temperature and aridity (annual heat to moisture index). In Figure 10, Omernik Level III Ecoregions (Omernik 1987) overlay the provisional seed zones to identify climatically similar but ecologically different areas. For site-specific disturbance regimes and restoration objectives, seed collection locations within a seed zone and ecoregion may be further limited by elevation, soil type, or other factors.
The Pacific Northwest Research Station’s (USDA FS PNW 2026) Threat and Resource Mapping (TRM) Seed Zone application provides links to interactive mapping features useful for seed collection and deployment planning. The Climate Smart Restoration Tool (Richardson et al. 2020) can also guide revegetation planning, seed collection, and seed deployment, particularly when addressing climate change considerations.
Occurrence Map
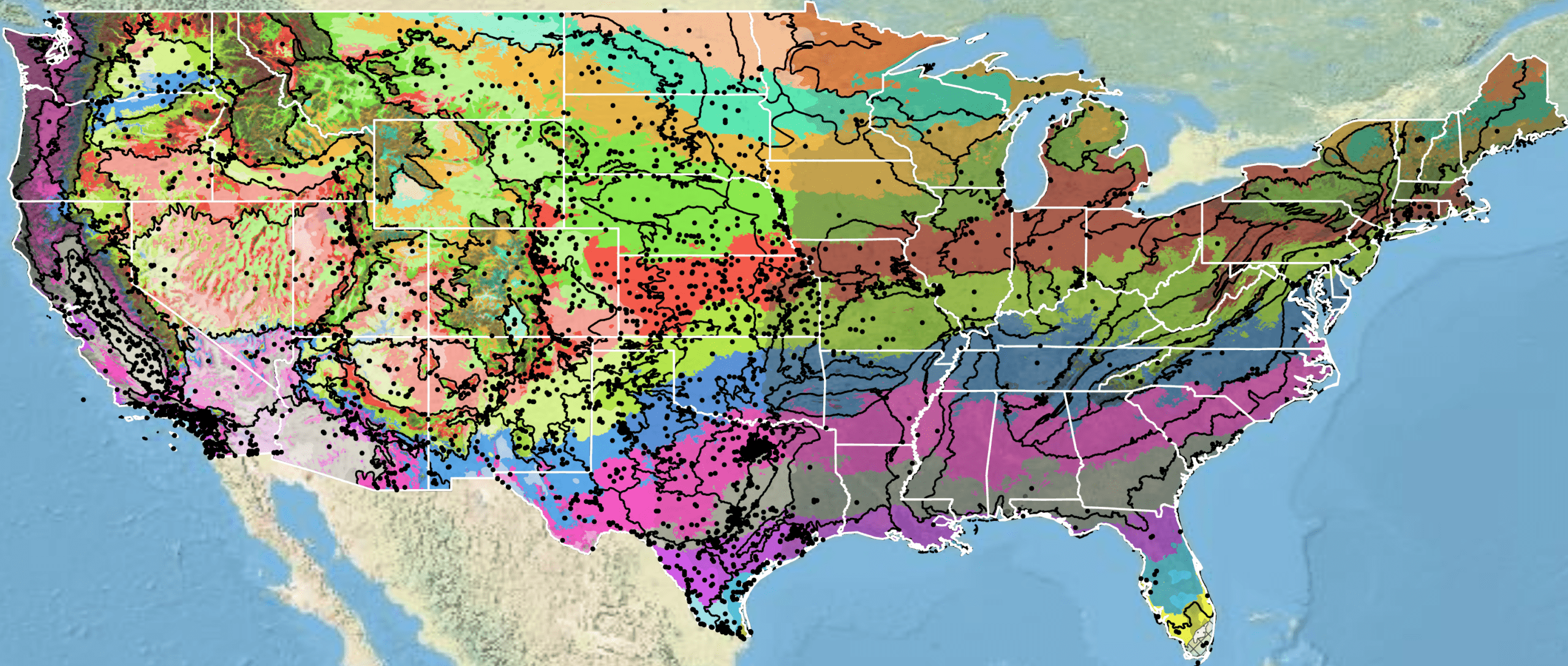
Figure 10. Distribution of common sunflower (black circles) based on geo-referenced herbarium specimens and observational data from 1828-2019 (CPNWH 2020; SEINet 2020; USDI USGS 2020). Generalized provisional seed zones (colored regions) (Bower et al. 2014) are overlain by Omernik Level III Ecoregions (black outlines) (Omernik 1987; USDI EPA 2018). Interactive maps, legends, and a mobile app are available (USDA FS PNW 2026; https://research.fs.usda.gov/pnw/products/dataandtools/seed-zone-webmap). Map prepared by Sarah Barga and Benton Spendlove USFS RMRS.
Releases
As of 2026, there were no common sunflower germplasm releases made for wildland restoration although there have been many cultivar releases for other uses.
Wildland Seed Collection
Common sunflower seeds are typically mature 49 to 56 days after flowering (Redente et al. 1982) and can be harvested as soon as flower petals shrivel (Wall and MacDonald 2009).
Wildland Seed Certification
Verification of species and tracking of geographic source is necessary whether wildland seed is collected for immediate project use or as stock seed for cultivated increase. This official Source Identification process can be accomplished by following procedures established by the Association of Official Seed Certifying Agencies (AOSCA) Pre-Variety Germplasm Program (UCIA 2015; Young et al. 2020). Wildland seed collectors should become acquainted with state certification agency procedures, regulations, and deadlines in the states where they collect.
If wildland-collected seed is to be sold for direct use in ecological restoration projects, collectors must apply for Source-Identified certification prior to making collections. Pre-collection applications, site inspections, and species and seed amount verification are handled by the AOSCA member state agency where seed collections will be made (see listings at AOSCA.org).
If wildland seed collected by a grower or private collector is to be used as stock seed for planting cultivated seed fields or for nursery propagation (See Agricultural Seed Certification section), detailed information regarding the collection site and collecting procedures must be provided when applying for certification. Photos and herbarium specimens may be required. Germplasm accessions acquired within established protocols of recognized public agencies, however, are normally eligible to enter the certification process as stock seed without routine certification agency site inspections. For contract grow-outs, however, this collection site information must be provided to the grower to enable certification.
Collection Timing
Common sunflower seeds can be harvested as soon as petals shrivel (Wall and MacDonald 2009). This timeframe can range from mid-June to October based on data from U.S. Department of Interior, Bureau of Land Management’s Seed of Success (SOS) collection crews (USDI BLM SOS 2024). The SOS teams made 47 collections from 2003 to 2019 in Oregon, California, Nevada, Arizona, New Mexico, Texas, Utah, Colorado, and Wyoming. Although collection dates ranged from June to October, most collections were made in August (11), September (16), and October (14). In the single year (2016) with the most collections (11) the earliest harvest date was August 18 at an elevation of 5,721 ft (1,744 m) in Utah and the latest was on October 5 at 5,378 ft (1,639 m) elevation also in Utah (USDI BLM SOS 2024).
Collection Methods
Common sunflower seed can be hand collected by clipping flower heads into a container (Fig. 11). Mature seeds will be released over time. Crushing or rubbing the flower heads to force the seeds from the head is not recommended since receptacle bracts remain attached and are difficult to remove (Wall and MacDonald 2009). Seed can also be collected by beating heads over the container. Reel-type harvesters and combines can be used in large natural stands or seed production plots (Jorgensen and Stevens 2004).
Based on seed collection in Nevada and Utah, the plant resource pursuit rate was 5.8 to 6.5 hrs/kg, processing rate was 0.7 to 2.0 hrs/kg for a return rate of 467 to 504 calories/hr (Simms 1985).

Figure 11. Collection of hundreds of dry common sunflower seed heads in a paper bag. Photo: USDI Bureau of Land Management (BLM) UT 020 Seeds of Success (SOS).
Several collection guidelines and methods should be followed to maximize the genetic diversity of wildland collections: 1) collect seed from a minimum of 50 randomly selected plants; 2) collect from widely separated individuals throughout a population without favoring the most robust or avoiding small stature plants; and 3) collect from all microsites including habitat edges (Basey et al. 2015). General collecting recommendations and guidelines are provided in online manuals (e.g., ENSCONET 2009; USDI BLM SOS 2021).
It is critical that wildland seed collection does not impact the sustainability of native plant populations. Collectors should take no more than 20% of the viable seed available at the time of harvest (USDI BLM SOS 2021). Additionally, care must be taken to avoid the inadvertent collection of crop or crop-wild hybrid types (Fig. 12).

Figure 12. Common sunflower seeds measuring about 0.5 in (5 cm) long and half as wide. Photo: USDI Bureau of Land Management (BLM) CA 180 Seeds of Success (SOS).
Post-Collection Management
Flower heads and seed should be kept in a dry, shaded place until collections can be moved to a controlled short-term storage environment. Short-term storage should be dry, cool, and inaccessible to rodents or other seed predators. If insects are suspected in any collection, seed should be frozen for 48 hours or treated with an appropriate insecticide. The more plant material in the collection, the more ventilation and drying a seed lot will likely need (Gold n.d.; Hay and Probert 2011).
Seed Cleaning
Few details about cleaning common sunflower seed were reported in the reviewed literature. Jorgensen and Stevens (2004) suggest processing harvested seedheads through a hammermill followed with a barley debearder, and then an air screen separator and gravity table.

Figure 13. Common sunflower seed under magnification. Photo: USDI Bureau of Land Management (BLM) CA 180 Seeds of Success (SOS).
Seed Storage
Although some suggest common sunflower seed is short lived (< 5 years in storage) (Baskin and Baskin 1998), but a study in Texas found that seed stored in a jar in a laboratory for 20 years was 90% viable (Seiler 2010). The wild seed was collected near Vega, Texas and stored in a glass jar at room temperature (68-72 °F [20-22 °C]) in a laboratory. Treating stored seeds with 1 mM GA3 increased germination from 13%. to 89% (Seiler 2010). The same seed and procedures revealed that 44.6% of seed was still viable after 38 years of room temperature storage. Germination of untreated long-stored seed was 5.5% but increased to 38.2% with GA3 treatment (Seiler 2022).
Seed Testing
The Association of Official Seed Analysts provides procedures for testing seed viability (AOSA 2010) and testing germination (AOSA 2016) of common sunflower.
Viability Testing
The Association of Official Seed Analysts provide several preparation methods to evaluate or determine common sunflower seed viability (AOSA 2010). Seed is imbibed on media moistened with distilled water overnight at 68 to 77 °F (20-25 °C). Prepared seeds are then soaked in a tetrazolium chloride (TZ) solution as indicated in Table 2.
Table 2. Preparation procedures used in tetrazolium chloride (TZ) testing of common sunflower seed. Imbibed seeds are prepared by one of the three methods listed below.
| Method | TZ Conc (%) | Time (hr) | Temp (°F) |
| Cut seed laterally and remove distal end of cotyledons | 1.0 | Overnight to 24 | 86-95 |
| Cut seed longitudinally, leaving seed intact at distal end | 0.1 | 6 to overnight | 86-95 |
| Cut seed longitudinally, leaving seed intact at basal end | 0.1 | 6 to overnight | 86-95 |
Evaluation of treated seeds (AOSA 2010):
After preparation, seed is considered viable if the
- entire embryo is evenly stained, even if there is slight damage to the radicle tip,
- radicle tip stains slightly darker or lighter than the rest of embryo
Seed is nonviable if:
- radicle with unstained areas extends into conducting tissue,
- cotyledons are half or more unstained or unstained near the point of attachment,
- any essential part of embryo is unstained,
- radicle has any unstained areas,
- there is extensive bruising (dark red, grey color),
- embryo is broken or cracked
Robinson et al. (2024) provide an alternate method for assessing the viability of common sunflower seed. Seeds are soaked in 1% TZ for 48 hours, then sliced through the pericarp, seed coat, and embryo laterally or diagonally at the wider, cotyledon end. The cut portion can be discarded. Forceps are then used to push the embryo out of the pericarp. Viable embryos stain pink (Robinson et al. 2024).
Germination Testing
The AOSA test for germination of common sunflower seed prescribes incubating seed on moist blotter paper, sand, or paper towel at a temperature of 68 °F (20 °C). The first count is made at 4 days and last at 7 days (AOSA 2016).
Germination Biology
Common sunflower seed matures in the fall and is dormant or conditionally dormant. Exposure to cold temperatures over winter reduce dormancy to enable spring germination (Baskin and Baskin 1998). Artificial cold stratification will encourage germination (Tolstead 1941a; Baskin and Baskin 1998; Buhler and Hoffman 1999).
Several researchers indicate that both afterripening (1-2 mos.) and cold exposure will maximize germination (Redente et al. 1982; Stevens et al. 1996). Germination of common sunflower seed wild collected in Garfield County, Colorado was 40% after exposure to 12 weeks of winter temperatures and less with exposure to temperatures that mimicked early to late spring temperatures. Seed treatments were conducted with 12 hr light/12 hr dark cycles (Kramer and Foxx 2016). Sheley and Half (2006) examined a seed lot with 59% viability and found that 22% found that 22% of common sunflower seed germinated readily and 33% was dormant. Seed was germinated on filter paper in 8 hr (77 °F [25 °C]) light/16 hr (59 °F [15 °C]) dark temperature cycles. Although the seed source was not identified, the project was focused on native seed (Sheley and Half 2006).
Germination of wild, crop-wild hybrids, and crop common sunflower seeds differ. Cultivated seed germinates readily to almost 100%, while wild seed germinates irregularly and is improved when placed outdoors for 3 to 4 weeks of alternating freezing and thawing temperatures (Heiser et al. 1969; Mercer et al. 2006). After being outdoors, wild seed moved into the greenhouse germinated at 50% or less (Heiser et al. 1969). Mercer et al. (2006) evaluated germination of wild and F1 crop-wild hybrids. Under laboratory conditions, wild seed germinated at 34% while hybrids germinated at 65% with a greater proportion of wild seeds (58%) remaining dormant than hybrid seeds (27%). Results were similar in a field study of germination where 35% of wild and 73% of hybrids germinated. Wild seed was collected from populations in Washington, Idaho, Montana, Wyoming, the Dakotas, Iowa, and Minnesota. Wild seed from Washington had high germination (82%) in the laboratory. In the laboratory study, seeds were cold stratified 39 °F (4 °C) for 25 days then exposed to 12 hr 77 °F (25 °C) light/12 hr (59 °F [15 °C]) dark cycles and monitored for germination for 14 days. Seed produced by plants grown from wild Idaho seed germinated at 43% in the field. In the field study, seeds were buried in metal mesh packets beneath 5 in (12.7 cm) of soil at the University of Minnesota Experimental Station in November and recovered at intervals from 10 April to 23 June (Mercer et al. 2006).
A low percentage of common sunflower seed germinated (<12%) after being submerged in water for 48 months but none germinated after 60 months of submergence. Seeds were collected from fields so may be crop or crop-wild hybrids. Seeds were submerged in a canal in Prosser, Washington beginning on July 1, 1963 (Comes et al. 1978).
Wildland Seed Yield And Quality
Post-cleaning seed yield and quality of seed lots collected in the Intermountain region are provided in Table 3 (USDA FS BSE 2025). The results indicate that common sunflower seed can generally be cleaned to high levels of purity and seed fill and viability of fresh seed can be variable.
Table 3. Seed yield and quality of common sunflower seed lots collected in the Intermountain region, cleaned by the Bend Seed Extractory, and tested by the Oregon State Seed Laboratory or the USDA Forest Service National Seed Laboratory (USDA FS BSE 2025).
| Seed lot characteristic | Mean | Range | Samples (no.) |
| Bulk weight (lbs) | 4.1 | 0.37-29 | 45 |
| Clean weight (lbs) | 1.1 | 0.04-5.8 | 45 |
| Clean-out ratio | 0.3 | 0.03-0.8 | 45 |
| Purity (%) | 98 | 95-99 | 45 |
| Fill (%)¹ | 92 | 73-99 | 45 |
| Viability (%)² | 94 | 68-99 | 43 |
| Seeds/lb | 73,226 | 56,770-98,780 | 45 |
| Pure live seeds/lb | 67,759 | 51,379-87,272 | 43 |
¹ 100 seed X-ray test
² Tetrazolium chloride test
Marketing Standards
Acceptable seed purity, viability, and germination specifications vary with revegetation plans. Purity needs are highest for precision seeding equipment used in nurseries, while some rangeland seeding equipment handles less clean seed quite well. Jorgensen and Stevens (2004) suggest that 90% purity and 70% germination are standard for common sunflower seed.
Agricultural Seed Production
Common sunflower was grown successfully for seed production by the Great Basin Research Center (Landeen and Gunnell 2018). Five accessions were grown in 85 ft x 5 ft (26 m x 1.5 m) plots at Fountain Green, Utah. Peek bloom date was August 31 to September 11. Seed was harvested on October 12. The amount of clean seed produced ranged from 1 to 2 lbs (480-880 g)/plot based on seed origin in plots that were 85 × 5 ft (26 × 1.5 m). The research center distributed 8.8 lbs (3,992 g) of common sunflower seed to other native forb seed growers in Oregon and Utah (Landeen and Gunnell 2018).
Agricultural Seed Certification
In order to minimize genetic changes in specific accessions of native species when increased in cultivated fields, it is essential to track the geographic source and prevent inadvertent hybridization or selection pressure. This is accomplished by following third party seed certification protocols for Pre-Variety Germplasm (PVG) as established by the Association of Official Seed Certification Agencies (AOSCA). AOSCA members in the U.S., Canada, and other countries administer PVG requirements and standards that track the source and generation of planting stock. Field and cleaning facility inspections then monitor stand establishment, proper isolation distances, control of prohibited weeds, seed harvesting, cleaning, sampling, testing, and labeling for commercial sales (UCIA 2015; Young et al. 2020).
Seed growers apply for certification of their production fields prior to planting and plant only certified stock seed of an allowed generation (usually less than four). The systematic and sequential tracking through the certification process requires preplanning, knowing state regulations and deadlines, and is most smoothly navigated by working closely with state certification agency personnel. See the Wildland Seed Certification section for more information on stock seed sourcing.
Weed Management
Mechanical and herbicide fallow is recommended to control weeds before planting common sunflower for seed production with cultivation of weeds as needed in the early part of growing season (Stevens et al. 1996).
Seeding
Fall seeding into soils ranging from sands to clay loams is recommended for common sunflower seed production. Drill seeding at depths of 0.1 to 0.5 in (0.2-1.3 cm) deep at a rate of 5 to 10 PLS/linear ft (32 PLS/m) of row with 24 in (61 cm) row spacing is also recommended (Stevens et al. 1996).
Irrigation
Little to no irrigation was required for common sunflower seed production at sites in Utah receiving 10 to 12 in (250-300 mm) of annual precipitation (Stevens et al. 1996).
Pollinator Management
The cavity-nesting Asteraceae specialist (Osmia californica) and the European honey bee (Apis mellifera) (Cane et al. 2012) can be transported to seed field production plots in portable ground nests or hives. If introducing new bee populations is not feasible, consider methods for encouraging and sustaining native bee populations where present to improve seed production (Cane 2008).
Seed Harvesting
As an annual, common sunflower stands produce one seed crop. Seed retention is considered fair, and seed can be harvested mechanically with a combine or other reel-type harvester (Stevens et al. 1996; Jorgensen and Stevens 2004).
Seed Yields And Stand Life
Stevens et al. (1996) report that common sunflower seed production yields of 300 to 450 lbs/ac (340-500 kg/ha) at 95% purity are possible.
Nursery Practice
Production of common sunflower nursery stock for revegetation was not reported in the reviewed literature.
Wildland Seeding And Planting
Common sunflower has many useful traits for revegetation and reclamation. Rapid growth rate, tolerance of a variety of soil types, importance to wildlife, and provision of summer and fall pollinator habitat have made it a common restoration species (Ogle et al. 2013, 2019). Recommended seeding rates of 13 to 30 PLS lbs/ac (14-34 PLS kg/ha) for sites receiving 6 to 16 in (152-406 mm) of annual precipitation are typical (Ogle et al. 2013, 2019).
Studies suggest common sunflower is useful in restoring burned, weedy, and mined sites. In a revegetation study of burned sagebrush and pinyon-juniper sites in west-central Utah, common sunflower established, although it was not part of the restoration seed mix (Ott et al. 2003). Sites were aerially seeded with primarily nonnative grasses following summer wildfires. A portion of seeded plots were chained to improve seed soil contact while the others were not. Common sunflower appeared at all sites for 3 years following the disturbance and restoration efforts (Ott et al. 2003).
Several studies suggest common sunflower may be useful for restoring weedy sites (Perry et al. 2009; Prasser and Hild 2016). In a greenhouse study, common sunflower grew well in the presence of competition with halogeton (Halogeton glomeratus) (Prasser and Hild 2016). Common sunflower seed was obtained from a commercial source was grown in pots (6 in [15 cm] diameter) filled with sandy clay loam as four conspecific seedlings or with two common sunflower seedlings and two halogeton seedlings. Pots were kept in a greenhouse with 12 hr (68 °F/20 °C) light/ 12 hr (50 °F/10 °C) dark cycles. After 14 weeks of growth, common sunflower was significantly taller (P = 0.0035) when grown with halogeton than in a monoculture. Halogeton biomass when grown with common sunflower was 11% of the total biomass. Survival of halogeton when grown with common sunflower was just 10% of that when grown with conspecifics (Prasser and Hild 2016).
A greenhouse experiment found that common sunflower reduced the cover and biomass of some nonnative species and increased the production of native species in an evaluation of its use as a cover crop (Perry et al. 2009). The study included nonnative species of cheatgrass (Bromus tectorum), Japanese brome (B. japonicus), Canada thistle (Cirsium arvense), and whitetop (Cardaria draba) and native species of hairy false goldenaster (Heterotheca villosa), green needlegrass (Nassella viridula), western wheatgrass (Pascopyrum smithii), and upright prairie coneflower (Ratibida columnifera). Seedlings of all species were grown for 30 to 38 days before being transplanted and utilized in addition treatments in the greenhouse with 8 h (68-82 °F [20-28 °C]) light and 16 h dark (55-68 °F [13-20 °C]) conditions. Pots were watered and fertilized primarily to limit competition for light. Common sunflower significantly reduced biomass of Japanese brome and cheatgrass (P < 0.0001) in the absence of native species and when grown with upright prairie coneflower, western wheatgrass, and hairy false goldenaster but not with green needlegrass. Common sunflower also significantly (P = 0.0002) reduced biomass of Canada thistle in the absence of native species and when grown with hairy false goldenaster or western wheatgrass but not with needlegrass or upright prairie coneflower. Common sunflower significantly reduced whitetop biomass (P < 0.0001) in the absence of native species or with hairy false goldenaster but not with needlegrass, wheatgrass, or upright prairie coneflower (Perry et al. 2009).
In a review of heavy metal pollution, common sunflower is considered a hyperaccumulator plant based on fast growth and maximum heavy accumulation efficiency (Jacob et al. 2018). It is also capable of phytostabilization by accumulating metals in its shoots and roots (Liu et al. 2024).
Common sunflower is recommended for reclamation of mine spoils because of its low pH tolerance, rapid establishment and growth, and soil stabilization. It is typically suggested for use with a mixture of species (Vogel 1981). Common sunflower is considered a non-invasive, rapid grower and has been used to restore mine sites in eastern Amazon, Brazil, and China (Gastauer et al. 2021; Meng et al. 2025).
Common sunflower naturally colonized several reclamation sites in the western U.S. (Holechek et al. 1981; Halvorson and Lang 1989). It colonized strip-mined land in southeastern Montana. Reclaimed sites were ripped, topsoiled, and broadcast seeded with a mix of native and nonnative species. Common sunflower occurred on the reclaimed sites in the first and fifth post reclamation years (Holechek et al. 1981). In soil contaminated during saltwater blowout used in oil drilling, common sunflower was a volunteer on sites treated with calcium chloride irrigation to leach sodium in 1982. The site was a Rocky Mountain juniper (Juniperus scopulorum)-grassland in the badlands of western North Dakota (mean annual ppt: 17 in [430 mm]). Following the irrigation treatment, the site was seeded with native grasses. Common sunflower occurred on nearby undisturbed plots and at a density of 0.8 plant/ft2 (9/m2) on non-reclaimed salt plots, and 0.5 plants/ft2 (5/m2) on reclaimed plots (Halvorson and Lang 1989).
Common sunflower also colonized a reclaimed surface mine site on the Missouri Plateau in western North Dakota. The mixed grass prairie reclamation following surface mining returned 5 to 8 in (13-20 cm) of topsoil from the native prairie that sat in piles for up to 2 years, included fertilization (200 lbs/ac [225 kg/ha] NPK), and was seeded with mostly nonnative grasses and forbs. Frequency of common sunflower was 18% in the first post reclamation year, 2% in the second (Iverson and Wali 1982).
Acknowledgements
Funding for Western Forbs: Biology, Ecology, and Use in Restoration was provided by the USDI BLM Great Basin Native Plant Materials Ecoregional Program through the Great Basin Fire Science Exchange. Great thanks to the chapter reviewers: Kimberly Wahl-Villarreal, DOI Bureau of Land Management and Nancy Jensen, USDA Natural Resources Conservation Service.
This research was supported in part by the USDA Forest Service, Rocky Mountain Research Station. The findings and conclusions in this publication are those of the authors and should not be construed to represent any official USDA or U.S. Government determination or policy.
Literature Cited
Albertson, F.W.; Weaver, J.E. 1944. Nature and degree of recovery of grassland from the great drought of 1933 to 1940. Ecological Monographs. 14(4): 394-479.
Alcoze, T.M.; Zimmerman, E.G. 1973. Food habitats and dietary overlap of two heteromyid rodents from the mesquite plains of Texas. Journal of Mammalogy. 54(4): 900-908.
Alexander, H.M., Cummings, C.L.; Kahn, L.; Snow, A.A. 2001. Seed size variation and predation of seed produced by wild and crop-wild sunflowers. American Journal of Botany. 88(4): 623-627.
Alexander, H.M.; Pilson, D.; Moody-Weis, J.; Slade, N.A. 2009. Geographic variation in dynamics of an annual plant with a seed bank. Journal of Ecology. 97(6): 1390-1400.
Alexander, H.M.; Schrag, A.M. 2003. Role of soil seed banks and newly dispersed seeds in population dynamics of the annual sunflower, Helianthus annuus. Journal of Ecology. 91(6): 987-998.
Andersen, B.A.; Holmgren, A.H. 1996. Mountain plants of northeastern Utah. HG 506. Logan, UT: Utah State University. 124 p.
Association of Official Seed Analysts [AOSA]. 2010. AOSA/SCST tetrazolium testing handbook. Contribution No. 29. Lincoln, NE: Association of Official Seed Analysts.
Association of Official Seed Analysts [AOSA]. 2016. AOSA rules for testing seeds. Vol. 1. Principles and procedures. Washington, DC: Association of Official Seed Analysts.
Bani-Yaghoub, M.; Reed, A. 2018. A methodology to quantify the long-term changes in social networks of competing species. Ecological Modelling. 368: 147-157.
Barney, M.A. 1972. Vegetation changes following fire in the pinyon-juniper type of west central Utah. Provo, UT: Brigham Young University. Thesis. 71 p.
Basey, A.C.; Fant, J.B.; Kramer, A.T. 2015. Producing native plant materials for restoration: 10 rules to collect and maintain genetic diversity. Native Plants Journal. 16(1): 37-53.
Baskin, C.C.; Baskin, J.M. 1988. Germination ecophysiology of herbaceous plant species in a temperate region. American Journal of Botany. 75(2): 286-305.
Baskin, C.C.; Baskin, J.M. 1998. Chapter 5: Germination biology of seeds with morphophysiological dormancy. In: Seeds: Ecology, biogeography, and evolution of dormancy and germination. San Diego, CA, Academic Press: 87-100.
Beatley, J.C. 1976. Vascular plants of the Nevada Test Site and central-southern Nevada: Ecologic and geographic distributions. TID-26881. Washington, DC: Technical Information Center, Energy Research and Development Administration. 306 p.
Bower, A.D.; St. Clair, J.B.; Erickson, V. 2014. Generalized provisional seed zones for native plants. Ecological Applications. 24(5): 913-919.
Buhler, D.D.; Hoffman, M.L. 1999. Andersen’s guide to practical methods of propagating weeds and other plants. Lawrence, KS: Allen Press. 248 p.
Campbell, H.; Martin, D.K.; Ferkovich, P.E.; Harris, B.K. 1973. Effects of hunting and some other environmental factors on scaled quail in New Mexico. Wildlife Monographs. 34: 3-49.
Canadell, J.; Jackson R.B.; Ehleringer, J.R.; Mooney, H.A.; Sala, O.E.; Schulze, E.D. 1996. Maximum rooting depth of vegetation types at the global scale. Oecologia. 108: 583-595.
Cane, J.H. 2008. Pollinating bees crucial to farming wildflower seed for U.S. habitat restoration. In: James, R.; Pitts-Singer, T., eds. Bees in agricultural ecosystems. Oxford, UK: Oxford University Press: 48-64.
Cane, J.H. 2012. Pollination and breeding biology studies. In: Shaw, N.; Pellant, M., eds. Great Basin Native Plant Project: 2011 Progress Report. Boise, ID: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station: 173-176.
Cane, J.H.; Love, B.; Swoboda, K. 2012. Breeding biology and bee guild of Douglas’ dustymaiden, Chaenactis douglasii (Asteraceae, Helenieae). Western North American Naturalist. 72(4): 563-568.
Carney, S.E.; Gardner, K.A.; Rieseberg, L.H. 2000. Evolutionary changes over the fifty-year history of a hybrid population of sunflowers (Helianthus). Evolution. 54(2): 462-474.
Castetter, E.F.; Opler, M.E. 1936. Ethnobiological studies in the American Southwest III. The ethnobiology of the Chiricahua and Mescalero Apache. The University of New Mexico Bulletin. 4(5): 3-63.
Chamberlin, R.V. 1911. The ethno-botany of the Gosiute Indians of Utah, Memoirs of the American Anthropological Association. 2(5): 331-405.|
Chambers, K.L. 2020. Dicots: Asteraceae. In: Meyers, S.C.; Jaster, T; Mitchell, K.E.; Harvey, T.; Hardison, L.K., eds. Flora of Oregon. Volume 2: Dicots A-F. Fort Worth, TX: Botanical Research Institute of Texas (BRIT): 152-370.
Clary, W.P.; Jameson, D.A. 1981. Herbage production following tree and shrub removal in the pinyon-juniper type of Arizona. Journal of Range Management. 34(2): 109-113.
Comes, R.D., Bruns, V.F.; Kelley A.D. 1978. Longevity of certain weed and crop seeds in fresh water. Weed Science. 26(4): 336-344.
Consortium of Pacific Northwest Herbaria [CPNWH]. 2020. Seattle, WA: University of Washington Herbarium, Burke Museum of Natural History and Culture. pnwherbaria.org/
Costello, D.F. 1944. Natural revegetation of abandoned plowed land in the mixed prairie association of northeastern Colorado. Ecology. 25(3): 312-326.
Cottam, C. 1929. The status of the ring-necked pheasant in Utah. The Condor. 31(3): 117-123.
Craighead, J.J.; Craighead, F.C. Jr.; Davis, R.J. 1963. A field guide to Rocky Mountain wildflowers from northern Arizona and New Mexico to British Columbia. Boston, MA: Houghton Mifflin Company. 277 p.
Crawford, J.A.; Bolen, E.G. 1976. Fall diet of lesser prairie chickens in west Texas. The Condor. 78(1): 142-144.
Cronquist, A. 1994. Volume five: Asterales. In: Cronquist, A.; Holmgren, A.H.; Holmgren, N.H.; Reveal, J.L.; Holmgren, P.K., eds. Intermountain flora: Vascular plants of the Intermountain West, U.S.A. Bronx, NY: The New York Botanic Garden. 496 p.
Cummings, C.L.; Alexander, H.M. 2002. Population ecology of wild sunflowers: Effects of seed density and post-dispersal vertebrate seed predators. Oecologia. 130: 274-280.
Cummings, C.L.; Alexander, H.M.; Snow, A. A. 1999. Increased pre-dispersal seed predation in sunflower crop-wild hybrids. Oecologica. 121: 330-338.
Currah, R.; Smreciu, A.; Van Dyk, M. 1983. Prairie wildflowers: An illustrated manual of species suitable for cultivation and grassland restoration. Edmonton, AB: University of Alberta, Friends of the Devonian Botanic Garden. 290 p.
Daubenmire, R. 1975. Plant succession on abandoned fields, and fire influences, in a steppe area in southeastern Washington. Northwest Science. 49(1): 36-48.
Dechaine, J.M.; Burger J.C.; Burke, J.M. 2010. Ecological patterns and genetic analysis of post-dispersal seed predation in sunflower (Helianthus annuus) crop-wild hybrids. Molecular Ecology. 19: 3477-3488.
Dechaine, J.M.; Burger, J.C.; Chapman, M.A.; Seiler, G.J.; Brunick, R.; Knapp, S.J.; Burke, J.M. 2009. Fitness effects and genetic architecture of plant-herbivore interactions in sunflower crop-wild hybrids. New Phytologist.184: 828-841.
Discover Life. 2024. Sam Houston State University. Available: https://www.discoverlife.org/
Dorn, R.D.; Dorn, J.L. 2007. Growing native plants of the Rocky Mountain area. Morrisville, NC: Lulu.com. 260 p.
Ellstrand, N.C.; Torres, A.M.; Levin, D.A. 1978. Density and the rate of apparent outcrossing in Helianthus annuus (Asteraceae). Systematic Botany. 3(4): 403-407.
Elmore, F.H. 1944. Ethnobotany of the Navajo: A monograph of the University of New Mexico and the School of American Research No. 8. Santa Fe, NM: University of New Mexico. 136 p.
Erdman, J.A. 1970. Pinyon-juniper succession after natural fires on residual soils of Mesa Verde, Colorado. Brigham Young University Science Bulletin: Biological Series. 11(2): 1-26.
European Native Seed Conservation Network [ENSCONET]. 2009. ENSCONET seed collecting manual for wild species. Edition 1: 32 p.
Fautin, R.W. 1946. Biotic communities of the northern desert shrub biome in western Utah. Ecological Monographs. 16(4): 251-310.
Flinders, J.T.; Hansen, R.M. 1972. Diets and habitats of jackrabbits in northeastern Colorado. Range Science Department Science Series No. 12. Fort Collins, CO : Colorado State University. 29 p.
Gastauer, M.; Sanjuan de Medeiros Sarmento, P.; Frois Caldeira, C.; Flexa Castro, A.; Junio Ramos S.; Carreira Trevelin, L.; Jaffe, R.; Oliveira, G.; Walfir Martins Souza Filho, P. 2021. Shannon tree diversity is a surrogate for mineland rehabilitation status. Ecological Indicators. 130: 108100.
Gilmore, M.R. 1919. Uses of plants by the Indians of the Missouri River region. Lincoln, NE: University of Nebraska Press. 125 p.
Gold, K. n.d. Post-harvest handling of seed collections. Technical Information Sheet 04. UK: Royal Botanic Gardens Kew and Millennium Seed Bank Partnership. 4 p.
Goodrich, S.; Neese, E. 1986. Uinta Basin flora. Ogden, UT: U.S. Department of Agriculture, Forest Service, Intermountain Region. 320 p.
Grant-Hoffman, M.N.; Spencer A.; Parr, S.; Lincoln, A.; Sanders, B. 2003. Salt desert seeding trials to inform restoration seed mixes. Native Plants Journal. 24(3): 206-214.
Graves, S.D.; Shapiro, A.M. 2003. Exotics as host plants of the California butterfly fauna. Biological Conservation. 110: 413-433.
Halvorson, G.A.; Lang, K.J. 1989. Revegetation of a salt water blowout site. Journal of Range Management. 42(1): 61-65.
Harrington, H.D. 1967. Edible native plants of the Rocky Mountains. Albuquerque, NM: University of New Mexico Press. 392 p.
Hart, J. 1976. Montana native plants and early peoples. Helena, MT: Montana Historical Society Press. 152 p.
Hay F.R.; Probert, R.J. 2011. Chapter 20: Collecting and handling seeds in the field. In: Guarino, L.; Ramanatha, V.; Goldberg, E. Collecting plant genetic diversity: Technical Guidelines-2011 update. Rome, Italy: Bioversity International. 33 p.
Hazlett, D.L. 1998. Vascular plant species of the Pawnee National Grassland. Gen. Tech. Rep. RMRS-GTR-17. Fort Collins, CO, U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station: 26.
Heiser, C.B. 1947. Hybridization between the sunflower species Helianthus annuus and H. petiolaris. Evolution. 1(4): 249-262.
Heiser, C.B.; Smith D.M.; Clevenger, S.B.; Martin, W.C. 1969. The North American sunflowers (Helianthus). Memoirs of the Torrey Botanical Club. 22(3): 1-218.
Hermann, F. 1966. Notes on western range forbs: Cruciferae through Compositae. Agric. Handb. 293. Washington, DC: U.S. Department of Agriculture, Forest Service. 365 p.
Hickman, J.C., ed. 1993. The Jepson manual: Higher plants of California. Berkeley, CA: University of California Press. 1400 p.
Hitchcock, C.L.; Cronquist, A. 2018. Flora of the Pacific Northwest: An illustrated manual. Second Ed. Giblin, D.E.; Legler, B.S.; Zika, P.F.; Olmstead, R.G., eds. Seattle, WA: University of Washington Press. 882 p.
Holechek, J.L.; Depuit, E.J.; Coenenberg, J.G.; Valdez, R. 1981. Fertilizer effects on establishment of two seed mixtures on mined land in southeastern Montana. Journal of Soil and Water Conservation. 36(4): 241-244.
Iverson, L.R.; Wali, M.K. 1982. Reclamation of coal mined lands: The role of Kochia scoparia and other pioneers in early succession. Reclamation and Revegetation Research. 1: 123-160.
Jacob, J.M.; Karthik, C.; Ganesh Saratale, R.; Kumar, S.S.; Prabakar, D.; Kadirvelu, K.; Pagazhendhi, A. 2018. Biological approaches to tackle heavy metal pollution: A survey of literature. Journal of Environmental Management. 217: 56-70.
Johnson, J.R.; Larson, G.E. 1999. Grassland plants of South Dakota and the northern Great Plains. Research Bulletins of the South Dakota Agricultural Experiment Station. 566: 288.
Johnson, S.R.; Robel, R.J. 1968. Caloric values of seeds from four range sites in northeastern Kansas. Ecology. 49(5): 956-961.
Jorgensen, K.R.; Stevens, R. 2004. Seed collection, cleaning, and storage. In: Monsen, S.B.; Stevens, R.; Shaw, N.L., comps. Restoring western ranges and wildlands, vol. 3. Gen. Tech. Rep. RMRS-GTR-136-vol-3. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station: 699-716.
Kansas State University [KSU] n.d. Forage facts: Sunflower silage. FORA31. Manhattan, KS: Kansas State University Agricultural Experiment Station and Cooperative Extension Service. 2 p.
Karl, M.G.; Heitschmidt, R.K.; Haferkamp, M.R. 1999. Vegetation biomass dynamics and patterns of sexual reproduction in a northern mixed-grass prairie. The American Midland Naturalist. 141: 227-237.
Keil, D.J. 2020. Helianthus annuus. In: Jepson Flora Project (eds.) Jepson eFlora, Revision 8, https://ucjeps.berkeley.edu/eflora/eflora_display.php?tid=3244 [Accessed 2026 January 7].
Kelso, L. 1931. Some notes on young desert horned larks. The Condor. 33(2): 60-65.
Kindscher, K. 1987. Edible wild plants of the prairie: An ethnobotanical guide. Lawrence: University Press of Kansas. 279 p.
Knight, R.L.; Every, A.D.; Erickson, A.W. 1979. Seasonal food habits of four game bird species in Okanogan County, Washington. The Murrelet. 60(2): 58-66.
Kramer, A.; Foxx, A. 2016. Propagation protocol for production of propagules (seeds, cuttings, poles, etc.) Helianthus annuus seeds. Native Plant Network. U.S. Department of Agriculture, Forest Service, National Center for Reforestation, Nurseries, and Genetic Resources. http://npn.rngr.net/propagation/protocols [Accessed 2026 January 8].
Lady Bird Johnson Wildflower Center [LBJWC]. 2025. Helianthus annuus L. Native Plant Database. Austin, TX: Lady Bird Johnson Wildflower Center. https://www.wildflower.org/plants-main [Accessed 2026 September 13].
Landeen, M.; Gunnell, K. 2018. Great Basin Research Center Seed Increase. In: Kilkenny, F; Edwards, F.; Irwin, J.; Barga, S., eds. Great Basin Native Plant Project: 2017 Progress Report. Boise, ID: U.S. Department of Agriculture, Rocky Mountain Research Station: 90-95.
Lentz, D.L.; Pohl, M.E.; Pope, K.O.; Wyatt, A.R. 2001. Prehistoric sunflower (Helianthus annuus L.) domestication in Mexico. Economic Botany. 55(3): 370-376.
Lesica, P. 2012. Manual of Montana vascular plants. Fort Worth, TX: Botanical Research Institute of Texas (BRIT) Press. 779 p.
Liu, J.; Yao, J.; Tang, C.; Ma, B.; Liu, X.; Bashir, S.; Sunahara, G.; Duran, R. 2024. A critical review on bioremediation technologies of metal(loid) tailings: Practice and policy. Journal of Environmental Management. 359: 121003.
Livingston, R.B. 1949. An ecological study of the Black Forest, Colorado. Ecological Monographs. 19(2): 123-144.
Looman, J. 1963. Preliminary classification of grasslands in Saskatchewan. Ecology. 44(1): 15-29.
MacKinnon, A.; Kershaw, L.; Arnason, J.; Owen, P.; Karst, A.; Hamersley Chambers, F. 2014. Edible and medicinal plants of Canada. Edmonton, Alberta: Partners Publishing and Lone Pine Publishing. 448 p.
Martin, A.C.; Barkley, W.D. 2000. Seed identification manual. Caldwell, NJ: The Blackburn Press. 221 p.
Mayrose, M.; Kane, N.C.; Mayrose, I.; Dlugosch, K.M.; Rieseberg, L.H. 2011. Increased growth in sunflower correlates with reduced defenses and altered gene expression in response to biotic and abiotic stress. Molecular Ecology. 20: 4683-4694.
McKell, C.M. 1950. A study of plant succession in the oak brush (Quercus gambelii) zone after fire. Salt Lake City, UT: University of Utah. Thesis. 79 p.
Meng, D.; Qiu, J.; Huang, P.; Li, W.; Li, Z.; Li, H. 2025. Positive effects of no-till reseeding with Asteraceae plants on the remediation of arsenic-contaminated mining areas: Tolerance evaluation, community reconstruction, and soil health impacts. Journal of Environmental Management. 395: 128001.
Mercer, K.L., Shaw, R.G.; Wyse, D.L. 2006. Increased germination of diverse crop-wild hybrid sunflower seeds. Ecological Applications. 16(3): 845-854.
Michael, V.C.; Beckwith, S.L. 1955. Quail preference for seed of farm crops. The Journal of Wildlife Management. 19(2): 281-296.
Moerman, D. 2003. Native American ethnobotany: A database of foods, drugs, dyes, and fibers of Native American peoples, derived from plants. Dearborn, MI: University of Michigan. http://naeb.brit.org
Moody-Weis, J.; Alexander, H.M. 2007. The mechanisms and consequences of seed bank formation in wild sunflowers (Helianthus annuus). Journal of Ecology. 95: 851-864.
Mower, K.J.; Smith, H.D. 1989. Diet similarity between elk and deer in Utah. The Great Basin Naturalist. 49(4): 552-555.
Munz, P.A.; Keck, D.D. 1973. A California flora and supplement. Berkeley, CA: University of California Press. 1905 p.
Murie, A. 1946. The Merriam turkey on the San Carlos Indian Reservation. The Journal of Wildlife Management. 10(4): 329-333.
Ogle, D.; Pavek, P.; Fleenor, R.; Stannard, M.; Dring, T.; Cane, J.; Fullen, K.; St. John, L.; Tilley, D. 2013. Plants for pollinators in the Inland Northwest. Tech. Note 24. Boise, ID: U.S. Department of Agriculture, Natural Resources Conservation Service. 62 p.
Ogle, D.; Tilley, D.; Cane, J.; St. John, L.; Fullen, K.; Stannard, M.; Pavek, P. 2019. Plants for pollinators in the Intermountain West. Plant Materials Tech. Note 2A. Boise, ID: U.S. Department of Agriculture, Natural Resources Conservation Service. 40 p.
Omernik, J.M. 1987. Ecoregions of the conterminous United States. Map (scale 1:7,500,000). Annals of the Association of American Geographers. 77(1): 118-125.
Oregon State University. 2026. OregonFlora. Corvallis, OR: Department of Botany and Plant Pathology. https://oregonflora.org/taxa/garden.php?taxon=5524 [Accessed 2026 September 23].
Ostler, W.K.; Harper, K.T. 1978. Floral ecology in relation to plant species diversity in the Wasatch Mountains of Utah and Idaho. Ecology. 59(4): 848-861.
Ott, J.; McArthur; E.D.; Roundy, B. 2003. Vegetation of chained and non-chained seedings after wildfire in Utah. Journal of Range Management. 56: 81-91.
Ott, J.E.; McArthur, E.D.; Sanderson, S.C. 2001. Plant community dynamics of burned and unburned sagebrush and pinyon-juniper vegetation in west-central Utah. In: McArthur, E. D.; Fairbanks, D. J., comps. Shrubland ecosystem genetics and biodiversity: Proceedings; 2000 June 13-15; Provo, UT. Proc. RMRS-P-21. Ogden, UT: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station: 177-191.
Palmer, E. 1878. Plants used by the Indians of the United States. The American Naturalist. 12(9): 593-606.
Pavek, P.; Erhardt, B.; Heekin, T.; Old, R. 2012. Forb seedling identification guide for the Inland Northwest: Native, introduced, invasive, and noxious species. Pullman, WA: U.S. Department of Agriculture, Natural Resources Conservation Service, Pullman Plant Materials Center. 144 p.
Peck, M.E. 1911. Summer birds of Willow Creek Valley, Malheur County, Oregon. The Condor. 13(2): 63-69.
Pendleton, R.L.; Smith, B.N. 1983. Vesicular-arbuscular mycorrhizae of weedy and colonizer plant species at disturbed sites in Utah. Oecologia. 59(2/3): 296-301.
Perry, L.G.; Cronin, S.A.; Paschke, M.W. 2009. Native cover crops suppress exotic annuals and favor native perennials in a greenhouse competition experiment. Plant Ecology. 204(2): 247-259.
Peterson, J.G. 1969. The food habits and summer distribution of juvenile sage grouse in central Montana. Bozeman, MT: Montana State University. Thesis. 39 p.
Pieper, R.D.; Wittie, R.D. 1990. Fire effects in southwestern chaparral and pinyon-juniper vegetation. In: Krammes, J.S, Tech Coord. Effects of fire management of southwestern natural resources. Gen. Tech. Rep. RM-GTR-191. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Forest and Range Experiment Station: 87-93.
Pilson, D. 2000. Herbivory and natural selection on flowering phenology in wild sunflower, Helianthus annuus. Oecologia. 122: 72-82.
Plant Conservation Alliance [PCA]. 2015. National seed strategy for rehabilitation and restoration 2015-2020. Washington, DC: U.S. Department of the Interior, Bureau of Land Management. 52 p.
Prasser, N.P.; Hild, A.L. 2016. Competitive interactions between an exotic annual, Halogeton glomeratus, and 10 North American native species. Native Plants Journal. 17(3): 244-254.
Reagon, M. Snow, A.A. 2006. Cultivated Helianthus annus (Asteraceae) volunteers as a genetic bridge to weedy sunflower populations in North America. American Journal of Botany. 93(1): 127-133.
Redente, E.F.; Ogle, P.R.; Hargis, N.E. 1982. Growing Colorado plants from seed: A state of the art. Vol. III, forbs. FWS/OBS-82/30. Washington, D.C.: U.S. Department of the Interior, Fish and Wildlife Service. 141 p.
Reynolds, L.V.; Cooper, D.J. 2011. Ecosystem response to removal of exotic riparian shrubs and a transition to upland vegetation. Plant Ecology. 212(8): 1243-1261.
Richardson, B.; Kilkenny, F.; St. Clair, B.; Stevenson-Molnar, N. 2020. Climate Smart Restoration Tool. https://climaterestorationtool.org/csrt/
Richmond, T.D.A.; Mueller-Dombois, D. 1972. Coastline ecosystems on Oahu, Hawaii. Vegetatio 25(5/6): 367-400.
Rippel, P.; Pieper, R.D.; Lymbery, G.A. 1983. Vegetational evaluation of pinyon-juniper cabling in south-central New Mexico. Journal of Range Management. 36(1): 13-15.
Robinson, G.S.; Ackery, P.R.; Kitching, I.; Beccaloni, G.W.; Hernández, L.M. 2025. HOSTS: A Database of the World’s Lepidopteran Hostplants [Data set]. Natural History Museum. https://doi.org/10.5519/havt50xw
Robinson, R.; Johnston, S.E.; Feldman, E.; Houde, V.M.; Kurkowski, S.R.; Mast, A.; Braun, J.V.; Kettenring, K.M. 2024. Seed viability testing guide for common wetland plant species. Logan, UT: Utah State University Extension. 62 p.
Sandoval, L.; Holechek, J.; Biggs, J.; Valdez, R.; VanLeeuwen, D. 2005. Elk and mule deer diets in north-central New Mexico. Rangeland Ecology and Management. 58(4): 366-372.
Schilling, E.E. 2006. Helianthus L. Flora of North America North of Mexico Vol. 21 Magnoliophyta: Asteridae (in part). Flora of North America Editorial Committee. New York, NY: Oxford University Press. 21: 141-149.
Seiler, G.J. 2010. Germination and viability of wild sunflower species achenes stored at room temperature for 20 years. Seed Science and Technology. 38: 786-791.
Seiler, G.J. 2022. Germination and viability of wild sunflower species seeds stored at room temperature and low humidity for 38 years. Seed Science and Technology. 50(3): 307-315.
Seiler, G.J.; Qi, L.L.; Marek, L.F. 2017. Utilization of sunflower crop wild relatives for cultivated sunflower improvement. Crop Science. 57(3): 1039-1821.
SEINet – Regional Networks of North American Herbaria Steering Committee [SEINet]. 2020. SEINet Regional Networks of North American Herbaria. https://Symbiota.org/docs/seinet
Shay, T.C. 1986. Plants and people: past ethnobotany of the northeastern prairie. In: Clambey, G.K.; Pemble, R.H., eds. The prairie: past, present and future: Proceedings of the 9th North American Prairie Conference; 1984 July 29 – August 1; Moorhead, MN. Fargo, ND: Tri-College University Center for Environmental Studies: 1-7.
Sheley, R.L.; Half, M.L. 2006. Enhancing native forb establishment and persistence using a rich seed mixture. Restoration Ecology. 14(4): 627-635.
Shiflet, T.N. 1994. Rangeland cover types of the United States. Denver, CO: Society for Range Management. 152 p.
Simms, S.R. 1985. Acquisition cost and nutritional data on Great Basin resources. Journal of California and Great Basin Anthropology. 7(1): 117-126.
Skinner, M.P. 1928. The elk situation. Journal of Mammalogy. 9(4): 309-317.
Society for Ecological Restoration, International Network for Seed-based Restoration, and Royal Botanic Gardens Kew [SER, INSR, RBGK]. 2026. Seed Information Database (SID). Version 7.1. https://ser-sid.org/
Spellenberg, R. 2001. National Audubon Society field guide to North American wildflowers: Western region, revised edition. New York, NY: Alfred A. Knopf, Inc. 862 p.
Spellenberg, R. 2012. Sonoran Desert wildflowers: A guide to common plants. Helena, MT: Falcon Guides. 245 p.
Stephenson, T.E., Holecheck, J.L.; Kuykendall, C.B. 1985b. Drought effect on pronghorn and other ungulate diets. The Journal of Wildlife Management. 49(1): 146-151.
Stephenson, T.E., Holechek, J.L.; Kuykendall, C.B. 1985a. Diets of four wild ungulates on winter range in northcentral New Mexico. The Southwestern Naturalist. 30(3): 437-441.
Stevens, M. 2006. Plant guide: Annual sunflower: Helianthus annuus L. Greensboro, NC: U.S. Department of Agriculture, Natural Resources Conservation Service, National Plant Data Center. 5 p.
Stevens, O.A. 1932. The number and weight of seeds produced by weeds. American Journal of Botany. 19(9): 784-794.
Stevens, R.J.; Jorgensen, K.R.; Young, A.S.; Monsen, S.B. 1996. Forb and shrub seed production guide for Utah. AG 501. Logan, UT: Utah State University Extension. 52 p.
Strahan, R.T.; Stoddard, M.T.; Springer, J.D.; Huffman, D.W. 2015. Increasing weight of evidence that thinning and burning treatments help restore understory plant communities in ponderosa pine forests. Forest Ecology and Management. 353(1): 208-220.
Sullivan, T.P.; Sullivan, D.S.; Sullivan, J.H. 2017. Mammalian responses to windrows of woody debris on clearcuts: Abundance and diversity of forest-floor small mammals and presence of small mustelids. Forest Ecology and Management. 399: 143-154.
Teo-Sherrell, C.P.A. 1996. The fates of weed seeds. Lincoln, NE: University of Nebraska. Dissertation. 172 p.
Thacker, E.T.; Springer, T.L. 2016. Preference of pen-reared northern bobwhite among native plant seeds of the sand sagebrush-mixed prairie. The Southwestern Naturalist. 61(4): 307-311.
Thorn, T.D.; Zwank, P.J. 1993. Foods of migrating cinnamon teal in central New Mexico. Journal of Field Ornithology. 64(4): 452-463.
Tilley, D.; Pickett, T. 2021. Germination response of curlycup gumweed seed to oxygenated water treatment. Native Plants Journal. 22(1): 4-12.
Tilley, D.; Spencer, J.; Cracroft, T.; Brazee, B.; Vaughan, M.; Cruz, J.K. 2019. Creation and management of Utah butterfly habitat. Tech. Note Plant Materials 73. Boise, ID: U.S. Department of Agriculture, Natural Resources Conservation Service. 20 p.
Tolstead, W.L. 1941a. Germination habits of certain sand hill plants in Nebraska. Ecology. 22(4): 393-397.
Tolstead, W.L. 1941b. Plant communities and secondary succession in south-central South Dakota. Ecology. 22(3): 322-328.
Toole, E.H.; Brown, E. 1946. Final results of the Duvel buried seed experiment. Journal of Agricultural Research. 72(6): 201-210.
Turner, N.J.; Thompson, L.C.; Thompson, M.T.; York, A.Z. 1990. Thompson ethnobotany: Knowledge and usage of plants by the Thompson Indians of British Columbia. Royal British Columbia Museum Memoir No. 3. Victoria, British Columbia, Canada: Royal British Columbia Museum. 335 p.
Ungar, I.A.; Hogan, W.; McClelland, M. 1969. Plant communities of saline soils at Lincoln, Nebraska. The American Midland Naturalist. 82(2): 564-577.
University of Arizona. 2026. EcoRestore Portal: Restore Arizona’s native plants. https://ecorestore.arizona.edu/plant-list [Accessed 2025 September 22].
USDA Forest Service, Bend Seed Extractory [USDA FS BSE]. 2025. Nursery Management Information System Version 4.1.11. Local Source Report 34-Source Received. Bend, OR: U.S. Department of Agriculture, Forest Service Bend Seed Extractory.
USDA Forest Service, Pacific Northwest Research Station [USDA FS PNW]. 2026. Seed Zone WebMap. Portland, OR: U.S. Department of Agriculture, Forest Service, Pacific Northwest Research Station. https://research.fs.usda.gov/pnw/products/dataandtools/seed-zone-webmap
USDA Natural Resources Conservation Service [USDA NRCS]. 2026. The PLANTS Database. Greensboro, NC: U.S. Department of Agriculture, Natural Resources Conservation Service, National Plant Data Team. https://plants.sc.egov.usda.gov/
USDI Bureau of Land Management, Seeds of Success [USDI BLM SOS]. 2023. Bureau of Land Management technical protocol for the collection, study, and conservation of seeds from native plant species for Seeds of Success. Washington, DC: U.S. Department of the Interior, Bureau of Land Management, Plant Conservation and Restoration Program. 45 p.
USDI Bureau of Land Management, Seeds of Success [USDI BLM SOS]. 2024. Seeds of Success collection data. Washington, DC: U.S. Department of the Interior, Bureau of Land Management, Plant Conservation and Restoration Program.
USDI Environmental Protection Agency [USDI EPA]. 2018. Ecoregions. Washington, DC: U.S. Environmental Protection Agency. https://www.epa.gov/eco-research/ecoregions
USDI Geological Survey [USDI USGS]. 2020. Biodiversity Information Serving Our Nation (BISON). U.S. Geological Survey. https://www.gbif.us/
Utah Crop Improvement Association [UCIA]. 2015. How to be a seed connoisseur. Logan, UT: UCIA, Utah Department of Agriculture and Food, Utah State University and Utah State Seed Laboratory. 16 p.
Utah State University. 2026. EcoRestore Utah Portal. https://extension.usu.edu/ecorestore/assessment-tool?list=
Vestal, P.A. 1952. Ethnobotany of the Ramah Navaho. Reports of the Ramah Project: No. 4. Papers of the Peabody Museum of American Archeology and Ethnology. Cambridge, MA: Harvard University. 40(4): 1-94.
Walker, R.B. 1954. The ecology of serpentine soils: II. Factors affecting plant growth on serpentine soils. Ecology. 35(2): 259-266.
Wall, M.; MacDonald, J. 2009. Processing seeds of California native plants for conservation, storage, and restoration. Claremont, CA: Rancho Santa Ana Botanic Garden. 216 p.
Weaver, J.E. 1968. Prairie plants and their environment: A fifty-year study in the Midwest. Lincoln, NE, University of Nebraska Press. 276 p.
Weber, W.A. 1976. Rocky Mountain flora. Boulder, CO: Colorado Associated University Press. 479 p.
Welsh, S.L.; Atwood, N.D.; Goodrich, S.; Higgins, L.C., eds. 2015. A Utah Flora. Fifth Edition, revised. Provo, UT: Brigham Young University. 990 p.
Wienk, C.L.; Sieg, C.H.; McPherson, G.R. 2004. Evaluating the role of cutting treatments, fire and soil seed banks in an experimental framework in ponderosa pine forests of the Black Hills, South Dakota. Forest Ecology and Management. 192: 375-393.
Wilson, G.W.T.; Hartnett, D.C. 1998. Interspecific variation in plant responses to mycorrhizal colonization in tallgrass prairie. American Journal of Botany. 85(12): 1732-1738.
Wood, K.N.; Guthery, F.S.; Koerth, N.E. 1986. Spring-summer nutrition and condition of northern bobwhites in south Texas. The Journal of Wildlife Management. 50(1): 84-88.
Wright, H.A.; Bailey, A.W. 1980. Fire ecology and prescribed burning in the Great Plains: A research review. Gen. Tech. Rep. INT-77. Ogden, UT: U.S. Department of Agriculture, Forest Service, Intermountain Forest and Range Experiment Station. 48 p.
Young, S.A.; Schrumpf, B.; Bouck, M.; Moore, M. 2020. How the Association of Official Seed Certifying Agencies (AOSCA) tracks wildland sourced seed and other plant propagating materials. Logan, UT: Utah State University, Utah Agricultural Experiment Station. 36 p.
How to Cite
Gucker, C.L.; Shaw, N.L. 2026. Common sunflower (Helianthus annuus L.). In: Gucker, C.L.; Shaw, N.L., eds. Western forbs: Biology, ecology, and use in restoration. Reno, NV: Great Basin Fire Science Exchange. Online: https://westernforbs.org/common-sunflower-helianthus-annuus