Authorship
Gucker, Corey; Shaw, Nancy
Publication Date
August 2018
Update
March 2025
Nomenclature
Erigeron speciosus (Lind.) DC., hereafter referred to as aspen fleabane, belongs to the Astereae tribe of the Asteraceae or aster family (Nesom 2006).
Family
Asteraceae – Aster family
Genus
Erigeron
Species
speciosus
NRCS Plant Code
ERSP4 (USDA NRCS 2025).
Subtaxa
Although Flora North America (Nesom 2006) does not recognize subtaxa of aspen fleabane, geographically distributed morphological variation ascribed to varieties speciosus and maranthus (Cronquist et al. 1994; Welsh et al. 2015; USDA NRCS 2025) and potential hybridization with E. trinervis are discussed. Genetic variation and differentiation within the central region of the species range are described by Richardson et al. (2022, see Seed Sourcing section).
Synonyms
Erigeron conspicuus Rydberg, E. macranthus Nuttall, E. speciosus var. conspicuus (Rydberg) Breitung, E. speciosus var. macranthus (Nuttall) Cronquist, E. subtrinervis Rydberg ex Porter & Britton subsp. conspicuus (Rydberg) Cronquist, E. subtrinervis var. conspicuus (Rydberg) Cronquist, Stenactis speciosa Lindley (Nesom 2006).
Common Names
Aspen fleabane, Oregon fleabane, Oregon wild-daisy, showy daisy, showy fleabane (USDA FS 1937; Nesom 2006; AOSA 2016; USDA NRCS 2025).
Chromosome Number
Chromosome number is 2n = 18 (Jones and Young 1983). Two potentially triploid samples were identified by Richardson et al. (2022) using flow cytometry to determine genome size.
Hybridization
Distributions of aspen fleabane and threenerve fleabane (E. subtrinervis) have considerable overlap, and although they are considered at least partially reproductively isolated, intermediate forms are common (Nesom 2006).
Distribution
Aspen fleabane is widely distributed throughout western North America. In Canada, it occurs in British Columbia and Alberta. In the United States, it occurs in western South Dakota and all states to its west except California (Nesom 2006). Aspen fleabane is particularly common in the Great Basin and Intermountain regions (Hermann 1966). There is a population in Baja California, Mexico, that is disjunct from the nearest southernmost populations in Arizona (Nesom 2006).
Plants tolerate cold temperatures during the growing season, but flowering may be reduced by growing season frosts (Caradonna and Bain 2016). In experiments using tissue collected from plants growing near the Rocky Mountain Biological Laboratory (RMBL) near Gothic, Colorado, frost damage to 10% of leaf tissue occurred at 22 °F (-5.4 °C) and to 90% at 12 °F (-10.5 °C). Flowers were significantly more frost sensitive than leaves (P < 0.0001) for this late-flowering species. Ten percent of flowers were damaged at 26 °F (-3.2 °C) and 90% at 20 °F (-6.7 °C) (Caradonna and Bain 2016).
Habitat And Plant Associations
Aspen fleabane is widely distributed in wet and dry habitats (Ellison 1954; Inouye 2008; Pavek et al. 2012) where annual precipitation averages between 16 to 25 inches (410-640 mm) (Tilley et al. 2019). Small colonies are common (Mee et al. 2003), and plants may live for decades (Inouye 2008). Plants require full sun and are common in upper elevation meadows, big sagebrush (Artemisia tridentata) communities, quaking aspen (Populus tremuloides) woodlands, and coniferous forests (Mansfield 2000; Nesom 2006; UW Ext. 2018).
Herbaceous communities. Aspen fleabane is found in dry to wet grasslands and meadows. It is common in the tall forb rangeland cover type, occurring between 6,300 and 9,900 ft (1,900–3,000 m) elevation on deep soils (>1.5 ft [0.5 m]) in Montana, Idaho, Wyoming, Utah, Nevada, and Colorado (Fig. 1) (Shiflet 1994). It is commonly associated with Thurber’s fescue (Festuca thurberi) grasslands and upper elevation herbaceous communities in good condition (Hermann 1966). Near Crested Butte, Colorado, aspen fleabane occurred in dry, rocky, and wet montane meadows (Inouye 2008). It was also common in Thurber’s fescue grasslands at 9,500 to 10,500 ft (2,900-3,200 m) where soils had high clay content (Fall 1992, 1997). In these grasslands, aspen fleabane constancy was 84% and cover was 3.7% between 8,500 and 12,500 ft (2,600–3,800 m) elevation on steep, south slopes (Langenheim 1962). In eastern Oregon and Washington, aspen fleabane was associated with greenleaf fescue (F. viridula) subalpine grasslands (Franklin and Dyrness 1973).

Figure 1. Aspen fleabane growing in a central Idaho meadow. Photo: USFS.
Shrubland and woodland communities. Aspen fleabane is commonly found with big sagebrush (Costello 1944; Reed 1952; Langenheim 1962), quaking aspen (Reed 1952; Langenheim 1962; Robberecht and Caldwell 1987; Mansfield 2000), bigtooth maple (Acer grandidentatum), and Gambel oak (Quercus gambelii) (Allman 1953; Robberecht and Caldwell 1987). Aspen fleabane was abundant in big sagebrush and hillside quaking aspen vegetation types in the Jackson Hole Wildlife Park in Teton County, Wyoming (Reed 1952). Hillside quaking aspen stands had relatively open canopies and occurred primarily on southern slopes. Aspen fleabane did not occur in quaking aspen stands on level terrain with dense shade and moist soils (Reed 1952). In the Crested Butte area of Gunnison County, Colorado, aspen fleabane constancy was 82% and average cover was 1.5% in big sagebrush communities, which were common on southern slopes at 8,500 to 9,500 ft (2,600–2,900 m) elevation. Aspen fleabane also occurred in quaking aspen woodlands but was much more common (100% constancy and 5% average cover) in younger woodlands with a Thurber’s fescue understory than in mature woodlands with a forb-dominated understory (Langenheim 1962). Aspen fleabane was noted in the understory of bigtooth maple thickets on eastern slopes near Logan, Utah (Robberecht and Caldwell 1987) and in bigtooth maple-Gambel oak vegetation in the Wasatch Mountains near Provo, Utah (Allman 1953). In the Rocky Mountain foothills in Alberta, aspen fleabane was much more frequent in quaking aspen stands that were adjacent to grasslands than those adjacent to shrublands (Hersperger and Forman 2003).
Forest communities. Aspen fleabane often occurs in openings and understories of pine (Pinus spp.), Douglas-fir (Pseudotsuga menziesii), and spruce-fir (Picea–Abies spp.) forests. In southern Alberta, aspen fleabane was a common associate of Douglas-fir forests on sandy-gravelly foothills or rocky ridges in the montane zone at 4,500 to 6,500 ft (1,400–2,000 m) elevation (Moss 1955). On the Wasatch Plateau, Utah, aspen fleabane occurred in Engelmann spruce-subalpine fir (P. engelmannii–A. lasiocarpa) forests without dense tree cover. Although aspen fleabane was more abundant on mesic sites, it also occupied dry, south slopes, with prevailing western winds where overstory shade was lacking (Ellison 1954). Constancy of aspen fleabane was 80% in bristlecone pine (Pinus aristata)/Thurber’s fescue vegetation at 9,500 to 10,200 ft (2,900–3,100 m) elevation in the Front, San Juan, and Mosquito Mountain ranges in southern Colorado (Ranne et al. 1997). In the Rincon Mountains of Pima County, Arizona, aspen fleabane occurred between 6,300 and 8,300 ft (1,900 and 2,500 m) elevation in rich soils or on rock slopes in pine-oak, pine, and mixed-conifer forests (Bowers and McLaughlin 1987).
Elevation
Aspen fleabane typically occurs at elevations of 2,000 to 11,220 ft (600–3,420 m) (Nesom 2006; Welsh et al. 2015). Its range in Utah is reported as 5,770 to 11,220 ft (1,760–3,420 m) (Welsh et al. 2015).
Soils
Aspen fleabane is found on dry to moist sites with well-drained sandy, gravelly, or clay-loam calcareous or non-calcareous soils (Allman 1953; Craighead et al. 1963; Neely and Barkworth 1984; Holiday 2000; Nesom 2006).
Sandy to clay loam to gravelly soil textures were reported in aspen fleabane habitats. In Tsegi Canyon, northeastern Arizona, aspen fleabane was associated with quaking aspen stands on sandy soils (Holiday 2000). In subalpine Thurber’s fescue grasslands in the White River National Forest of western Colorado, aspen fleabane made up 1 to 4% of the vegetation composition where soils were 18 to 48 in (46–122 cm) deep and averaged 32% sand, 46% silt, and 22% clay with 30% gravel (Klemmedson 1956). The species was also common (production of up to 39 lbs/ac [44 kg/ha]), in high-elevation, Letterman’s needlegrass (Achnatherum lettermanii) communities where soils averaged 17% sand, 35% silt, and 48% clay with 9.2% gravel and 54% total porosity. Soils contained 6% organic matter, 0.3% total nitrogen, 0.14% total phosphorus, and 9,400 ppm potassium on average (Laycock and Richardson 1975). In bigtooth maple-Gambel oak vegetation in Utah’s Wasatch Mountains where aspen fleabane was common, soils were classified as silt loams, loams, or clay loams (Allman 1953). Soil pH averaged 6.6 to 7.2 for the A to C horizons, and moisture-holding capacity averaged 78% for the A horizon and 49% for the B and C horizons. Soluble salts averaged 255 ppm in A, 305 ppm in B, and 310 ppm in C horizons (Allman 1953).
Aspen fleabane was reported on both calcareous and noncalcareous soils (Neely and Barkworth 1984). In big sagebrush in the Bear River Range of southeastern Utah, aspen fleabane was slightly more common on calcareous dolomite than on noncalcareous quartzite soils. Dolomite soils had significantly higher pH levels and calcium and magnesium contents than quartzite soils. Clay content was similar but sand content was significantly lower and silt content greater in dolomite than in quartzite soils (P < 0.01) (Neely and Barkworth 1984). In the Crested Butte area of west-central Colorado, aspen fleabane was common in big sagebrush on limestone or shale soils (Langenheim 1962).
Warming experiments showed that aspen fleabane, a shallow-rooted forb and dominant contributor to aboveground biomass in control plots, exhibited reduced growth and abundance and increased moisture stress in heated plots (Harte 2019; Loik and Harte 1997). When researchers compared artificially warmed and control plots at the RMBL, they found lower soil moisture, higher soil temperatures, earlier snowmelt, and higher nitrogen mineralization rates on warmed plots (Harte 2019; Loik and Harte 1997). Offsite research resulted in increased biomass and flowering of aspen fleabane with water addition (P = 0.03) but not with nitrogen (N) (P = 0.08) (de Valpine and Harte 2001).
Slominski et al. (2018) examined the effect of rooting depth and N-fixing ability on the growth and physiological responses of four forbs to N addition and soil moisture availability, factors presumed to affect future plant distributions. Aspen fleabane was included as a shallow-rooted, non-nitrogen fixing species. The research was conducted in a subalpine meadow at 10,300 ft (3,150 m) elevation near the RMBL in Colorado. Ten controls and 10 N-addition plants of each species were randomly selected across a natural soil moisture gradient. On four dates beginning 28 June, ammonium nitrate in 250 ml water was added at 7- to 10-day intervals to a 1.5 × 1.5 ft (0.5 × 0.5 m) area around each of the N-addition plants (total application 9 g N/m2). Data was collected between 16 June and 3 August 2009 (Slominski et al. 2018).
Growth (number of stems × average stem height) increased 1.3% for each 1% increase in soil moisture measured at 7.9 in (20 cm) depth (Slominski et al. 2018). N-addition plants were 14% larger than controls (P = 0.074) after one season, but the expected positive interaction between soil moisture and N addition did not occur (P = 0.794). As anticipated, the results indicated that soil moisture had a strong effect on this shallow-rooted species. The unexpected low response of growth to N addition might indicate a delay beyond one season or that resources might have been devoted to belowground growth (Slominski et al. 2018).
Photosynthetic capacity based on chlorophyll fluorescence was increased by N addition (P < 0.001) as expected, and there was a marginal soil moisture and measurement date interaction (P = 0.051) (Slominski et al. 2018). Nitrogen additions did not affect transpiration rate (P = 0.370). Transpiration rate did not change with soil moisture on July 2 but decreased with increasing soil moisture on July 24 (P = 0.050). There was a marginally significant interaction between soil moisture and N addition on floral display (maximum average number of open flowers) with N additions decreasing floral display by 5.3% per each 1% increase in soil moisture, with no change across moisture gradients for control plants. The response may have indicated resource uptake below ground or interactions with other environmental factors or functional traits (Slominski et al. 2018).
Description
Aspen fleabane is a long-lived perennial with a thick, branching fleshy to woody caudex, branched rhizomes, and a taproot and fibrous root system (Ellison 1954; Craighead et al. 1963; Paulsen 1970; Welsh 1983; Welsh et al. 2015; Caradonna and Bain 2016). Clusters of mostly erect leafy stems branching in the upper portion and ranging from 6 to 40 in (15–100 cm) tall develop from the caudex (Fig. 2) (Welsh 1983; Spellenberg 2001; Nesom 2006; LBJWC 2018). Plants growing in montane meadows in Gothic, Colorado, averaged 15 stems per plant (Pardee et al. 2018).

Figure 2. Aspen fleabane growing in Utah. Photo: USDI BLM UT933 SOS.
Stems generally are sparsely covered with long hairs but are hairless just below the inflorescence (Weber 1976; Nesom 2006; Pavek et al. 2012). Stem and basal leaves are oblanceolate, spatulate, elliptic, or oval (Nesom 2006). They are entire, generally with clasping to subclasping petioles, three main veins, and are glabrous, except for the margins, which are often fringed with hairs (Cronquist 1943; Craighead et al. 1963; Weber 1976; Pavek et al. 2012; Riser and Meyers 2020). Basal leaves range from 1 to 6 in (3–15 cm) long and 0.2 to 0.8 in (4–20 mm) wide and usually wither by time of flowering (Nesom 2006; Welsh et al. 2015). Stem leaves (0.8–4 in [2–11 cm] long, 0.2–1 in [0.5–2.8 cm] wide) are even-sized along the bottom half of the stem or largest at the midpoint or base of the stems (Cronquist 1943; Welsh 1983; Nesom 2006; Welsh et al. 2015). They are generally reduced upward (Mee et al. 2003).
Flower heads arranged in corymb-like inflorescences number 1 to 20 or more per stem (Welsh 1983; Welsh et al. 2015; Pardee et al. 2018; Riser and Meyers 2020). Involucres are hemispheric, 11 to 22 mm wide, glandular with a few long hairs and two to three series of bracts (Welsh et al. 2015; Riser and Meyers 2020). Within a series, the bracts are more or less equal and non-overlapping, acuminate, somewhat spreading, and may have an orange stripe or purplish tip (Spellenberg 2001; Welsh et al. 2015; Riser and Meyers 2020). Flower heads are up to 2 in (5 cm) wide (Fig.3) (Shaw 1995; LBJWC 2018). They are comprised of 75 to 150 purple, blue, pink, or less commonly white, pistillate, fertile ray flowers, each with a narrow ligule (7-13 mm long, 1 mm wide), which surround yellow to orange, bisexual, fertile disk flowers (Craighead et al. 1963; Weber 1976; Nesom 2006; Welsh et al. 2015; LBJWC 2018). Fruits are two- to four-nerved, oblong and compressed or flattened cypselas up to 1.8 mm long (Nesom 2006). Fruits are hairy with two tan, bristly, pappi, the inner with 20 to 30 bristles (Cronquist 1943; Welsh 1983; Nesom 2006; Welsh et al. 2015). Mature aspen fleabane fruits are dark brown cypselas, about 1 to 2 mm long (Luna 2005; Lesica 2012). Although not true achenes, they are often referred to as achenes or seeds (Hermann 1966; Luna 2005). Throughout this review, seed will be used to refer to the cypsela and the seed it contains.

Figure 3. Aspen fleabane flower. Photo: USDI BLM OR931 SOS.
Site conditions and weather can affect the appearance and size of aspen fleabane plants (Louda et al. 1987; Pardee et al. 2018). In the Elk Mountain Range near Crested Butte, Colorado, plants growing in full sun were smaller than those growing in shade. Plants in the sun averaged 13 in (32 cm) tall, while those in shade averaged 16 in (40 cm) tall (P = 0.02). Leaf length averaged 1.8 in (4.6 cm) in the sun and 2.1 in (5.3 cm) in shade (P = 0.03) (Louda et al. 1987). In montane meadows near the RMBL in Gothic, Colorado, experimentally produced frosts occurring when flower buds were first visible (mid- to late June) reduced plant height by 29% (P < 0.001) and flower head size by 7.8% (P = 0.03) compared to controls (Pardee et al. 2018).
Reproduction
Aspen fleabane reproduces primarily from seed. Plants also produce rhizomes, and colony formation has been noted (Mee et al. 2003). Reproductive potential varies with climate and may be impacted by predicted future changes in climate. When frost conditions were experimentally applied to plants near the RMBL when the first flower buds were visible (mid- to late June), the total number of aspen fleabane flowers produced was 40% less than produced by non-frosted controls (P = 0.01) (Pardee et al. 2018). In warming experiments at the same location, 40% more flowers were produced in warmed than control plots (P = 0.05) (Lambrecht et al. 2007). Flower size, however, was significantly reduced by warming (P < 0.001). Reproductive effort or the relative amount of available carbon allocated to reproduction was not different between warmed and control plots, but seed production per plant was significantly lower in warmed than control plots (P = 0.02) (Lambrecht et al. 2007).
Phenology
Aspen fleabane is a late-flowering species. Flowers appear from June to October (Paulsen 1970; Holiday 2000; Nesom 2006), and seed production generally follows flower production by 1 to 2 months (Paulsen 1970; West and Reese 1984).
Climate and site conditions influence the timing and speed of reproduction (Paulsen 1970; Weaver and Collins 1977; West and Reese 1991). Timing of flowering is likely closely linked to timing of snow melt. In subalpine vegetation in Ephraim Canyon, Utah, flowering began about 55 days after snow melt and continued for about 35 days (Ellison 1954). In subalpine meadows in the Wasatch Mountains of northern Utah, flowering began about 60 days after snow melt, and the period from flowering to plant senescence was about 2 months (West and Reese 1984). In bigtooth maple-Gambel oak vegetation in the Wasatch Mountains, aspen fleabane flowers were observed from early July through early September over an 0.8 ac (0.3 ha) area (Allman 1953). Based on 41 years of records, the average date for peak flowering in an alpine meadow near the RMBL was August 3 (Caradonna and Bain 2016). In Tsegi Canyon, northeastern Arizona, aspen fleabane flowered in June and July (Holiday 2000).
Phenology and flower production vary with site and weather conditions and can be delayed with shading, increased moisture, and decreased temperature. In Utah’s Bear River Range, peak flowering for aspen fleabane was 22 days later in aspen woodlands than in meadows (West and Reese 1991). In a high-elevation Thurber’s fescue meadow in southwestern Colorado, aspen fleabane flowered from about June 20 to August 5 and produced its first mature seeds near the end of July in the warmest, driest summer over a 4-year period. Flowering occurred from June 30 to August 20, and seed maturation began around August 20 in the coolest, wettest summer (Paulsen 1970).
In 30 years of monitoring dry and wet montane meadows at the RMBL, Inouye (2008) noted a strong relationship between snow melt dates and summer flowering abundance. Greater snowpack and later snowmelt were associated with later initiation of growth and lower flower bud mortality. Earliest flowering was July 9 and latest was August 17. If snow melted before May 19 or if snowpack was less than 3 feet (1 m) on April 30, frost damage and reduced flowering were likely. Flower production in each plot occurred on average in 22.3 years during the 30-year study (Inouye 2008).
Experiments also reveal links between phenology, weather, and climate conditions (Weaver and Collins 1977; Dunne et al. 2003). Aspen fleabane flowering was earliest in years with normal snowpack levels (2 ft [0.6 m]) and latest when 8-ft (2.4 m) snowpack levels were created with snow fences in meadows (Idaho fescue-bearded wheatgrass [F. idahoensis–Elymus caninus]) in the Bridger Mountains near Bozeman, Montana. Flowering duration was shortest with 4-ft (1.2 m) snowpack levels and longest with normal snowpack levels (Weaver and Collins 1977). At the RMBL, flowering was significantly earlier (P < 0.001) and longer (P = 0.051) on experimentally warmed than control plots. Earlier snow removal (conducted experimentally without warming) also resulted in significantly earlier flowering than on control plots (P = 0.017) (Dunne et al. 2003).
Research in subalpine meadows at the RMBL (9,500 ft [2,900 m]) found that winter temperatures increased, the snowpack decreased by 33%, and the timing of snowmelt advanced significantly from 1973 to 2016 (Wadgymar et al. 2018). Flowering of aspen fleabane advanced (P < 0.0001), but days from snowmelt to first flowering remained about the same for this species and other late-flowering species.
Breeding System
Aspen fleabane is weakly self-compatible, but insect pollination is important for good seed production (Pardee et al. 2018; Ingold et al. 2024).
In a reproductive ecology study of aspen fleabane in conducted in sublpine meadows at about 9,500 ft (2,900 m) at the Rocky Mountain Biological Laboratory in Colorado, open pollinated plants produced an average of 61.2 viable seeds per capitulum (Ingold et al. 2024). Production was similar to open pollination when plants were hand pollinated by near-outcrossing (plants <82 ft [25 m] apart; 54.2 seeds/capitulum) and far-outcrossing (plants 330-3,300 ft [100-1,000 m] apart; 41.1 seeds/capitulum). Plants in self-pollination treatments (autogamy in bagged capitula and artificial geitonogamy), however, produced significantly fewer viable seeds (<5 seeds/capitulum). Viable seeds per ovule varied across treatments (P < 0.0001). Greatest percentages of viable seeds were produced by ovules of the open pollinated (49.1%) and near-outcross plants (51.8%). The far-outcross treatment produced significantly fewer seeds per ovule (36.7%), and production of viable seeds per ovule (<7%) was significantly lower for the two self-pollination treatments than for the other three treatments, possibly indicating outbreeding depression. The results indicated that the species is primarily self-incompatible, though with weak self-compatibility and the location was not pollinator limited (Ingold et al. 2024).
Pollination
Aspen fleabane attracts and is important to a diversity of pollinating insects. Experiments at the RMBL highlighted the importance of pollinators for seed production in general and for seed production following sub-optimal weather conditions (Pardee et al. 2018). Frosts experimentally applied in mid- to late-June when the first aspen fleabane flower buds appeared resulted in 40% fewer total flowers (P = 0.01) and smaller flower heads (P < 0.01) and 48% fewer pollinator visits (P = 0.04) than on control plots. On frost plots, however, pollinators spent 33% more time visiting individual flowers. Seed production increased with total pollinator visits (P = 0.001), and this relationship was stronger for frost treatments, which suggested frost had a negative effect on plant reproduction, but increased duration of pollinator visits helped to partially, but not fully, salvage seed production (Pardee et al. 2018).
Aspen fleabane flowers are visited by and provide an important food source to a variety of insects. In a survey of montane bee fauna on the Tonasket Ranger District of the Okanogan-Wenatchee National Forest in northern Washington, 42 bee species were collected from aspen fleabane from June 28 to July 9 and August 9 to August 14. The following bees: bumble bees (Bombus bifarius, B. insularis, B. mixtus), leafcutter bees (Megachile relativa), long-horned bees (Melissodes microsticta, M. pallidisignata), and mason bees (Osmia coloradensis, O. subaustralis) were collected more than five times from aspen fleabane flowers (Wilson et al. 2010). In a study from 2005 to 2007 in meadows near the RMBL, Burkle and Irwin (2009) observed plant-pollinator interactions between aspen fleabane and the following insect families: Andrenidae, Anthomyiidae, Apidae, Bombyiidae, Braconidae, Calliphoridae, Cleridae, Colletidae, Halictidae, Hesperiidae, Ichneumonidae, Lycaenidae, Megachilidae, Muscidae, Noctuidae, Nymphalidae, Rhagionidae, Sarcophagidae, Sphecidae, Stratiomyidae, Syrphidae, and Tachinidae (Burkle and Irwin 2009; see also: Discover Life 2025; Robinson et al. 2025). In a mountain fescue (Festuca spp.) grassland in central Colorado, flowers were visited by Queen Alexandra’s sulphur butterflies (Colias alexandra) (Watt et al. 1974). In Glacier National Park, moths (Scrobipalpula erigeronella and Coleophora brunneipennis) were collected from aspen fleabane which was thought to be a larval food source (Braun 1921). The northern checkerspot butterfly (Chlosyne palla) uses aspen fleabane as a host plant (Scott cited in Wahlberg 2001).
In subalpine meadows near RMBL, aspen fleabane was the primary nectar source for the Mormon fritillary (Speyeria mormonia), likely because aspen fleabane flowering coincides with the peak male and female butterfly flight season (mid- to late-July and August). In years when there were 300 or more flowers in the survey plots (23 of 35 yrs), 70 to 90% of all female visits and 60 to 90% of all male visits were to aspen fleabane flowers (Boggs and Inouye 2012).
In the same general area, aspen fleabane was also important to bumble bees (Bombus spp.) Almost 80% of bee visits were from B. bifarius, B. occidentalis, and B. flavifrons (Pleasants 1980). Bumble bees spent an average of 0.8 second handling aspen fleabane flowers, 0.14 second traveling between flowers within an inflorescence. Net energy intake was 0.08 J/s, which ranked fourth among the nine species evaluated (Pleasants 1981).
Flower color and size of aspen fleabane and size factor into pollinator visitation (Pohl et al. 2011). In subalpine meadows near RMBL, blue copper butterflies (Lycaena heteronea) preferred the white flowers of common yarrow (Achillea millefolium var. alpicola) over purple aspen fleabane flowers (P < 0.05), even when aspen fleabane flowers were painted white (P < 0.05). When given a choice between large and small aspen fleabane flowers, blue copper butterflies preferred large flowers (P < 0.05). In contrast, ruddy copper butterflies (L. rubidus) preferred aspen fleabane over common yarrow (P < 0.0001), and small wood-nymphs (Cercyonis oetus) preferred large aspen fleabane flowers over small flowers (P < 0.002).
Aspen fleabane and 22 other species commonly used in seed mixes for revegetation projects in western Montana were assessed for their attractiveness and use by bees (Glenny et al. 2022). Pollinator-friendliness scorecards were used to rank species by visitation rate, number of bee species attracted, support of specialist bees, duration of the blooming period, and extent of the species distribution. Aspen fleabane ranked in the top 25th percentile for all categories except bloom duration for which it was in the bottom 25th percentile. Overall, aspen fleabane was ranked in the upper one-third of these species and was recommended for use in pollinator restoration on western Montana sites at low to middle elevations (<6,500 ft [2,000 m]) with low water availability, high noxious weed potential, and longer growing seasons. This includes sagebrush grasslands, limber pine, ponderosa pine, Douglas-fir, and some lodgepole pine communities (Glenny et al. 2022).
Characteristics of nine forb species used in wildflower strips to provide floral resources to wild bees in agroecosystems were examined on four farms over 2 years (2013-2014) in Montana to determine functional roles of each species in supporting wild bee communities (Burkle et al. 2020a). Bee abundance, richness, and composition varied among farms and years, but sticky purple geranium (Geranium viscosissimum) and aspen fleabane were spatially and temporally consistent in supporting the greatest richness of wild bees. Both were visited by generalist and specialist species (Burkle et al. 2020a).
Examination of volatile organic compound traits of 14 forbs in a subalpine Montana meadow at 5,200 to 5,700 ft (1600-1750 m) elevation over 3 years (2012, 2015, 2016) revealed that intraspecific variation in dispersion of floral volatile organic compounds varied among species, but not among years (P < 0.0023) (Burkle et al. 2020b). Dispersion was greatest for aspen fleabane, arrowleaf balsamroot, and yellow avalanche lily (Erythronium grandiflorum). These results may indicate stabilized selection from pollinators or rigid biochemical pathways. Forbs with high dispersion may attract generalist pollinators and a broader number of bee species thus improving the likelihood of pollination. This is important as pollinator community composition often varies among years (Burkle et al. 2020b).
Ecology
Aspen fleabane is a long-lived, somewhat shade tolerant species capable of colonizing early seral sites. It is also associated with communities in near pristine condition. It was noted in the early colonization of sites in primary succession (Ellison 1949; Langenheim 1956) and was common in pristine upper elevation herbaceous communities (Hermann 1966). In the subalpine zone of central Colorado, aspen fleabane occurred in early seral perennial forb and Thurber’s fescue communities present by 1947 after a 1923 landslide (Langenheim 1956). It was also noted in early surveys of patchy vegetation recovery on severely over-grazed and subsequently flooded sites on the Wasatch Plateau in central Utah. Aspen fleabane was most frequent within patches of established penstemon (Penstemon spp.) or yarrow (Achillea spp.), and once established, it spread quickly (Ellison 1949).
Although commonly found in both sun and shade conditions, aspen fleabane abundance is typically greater in sunny than shady sites. In the Elk Mountain Range near Crested Butte, Colorado, frequency of aspen fleabane was greater in sunny meadows than in shaded quaking aspen woodlands (1.2 sun:shade) (Louda et al. 1987). When effects of clearcutting were examined in Utah’s Fish Lake National Forest, researchers reported that aspen fleabane directly benefitted from quaking aspen removal (Mueggler and Bartos 1977).
Seed And Seedling Ecology
Aspen fleabane produces light-weight, wind-dispersed seeds (Luna 2005), but once deposited in the soil, their longevity is unknown. Aspen fleabane was often encountered in what were likely 1-year-old or current year’s seed banks (Willms and Quinton 1995; Reynolds and Cooper 2011).
Aspen fleabane germinated from soil samples collected in June from riparian sites in northeastern Arizona’s Canyon de Chelly National Monument (Reynolds and Cooper 2011). Although the exact June sampling date was not reported, soil samples likely captured banked seed, because earliest flowering occurs in June and seed maturation follows that by a month or more (see Phenology section).
In studies conducted by Wills and Quinton (1995) in rough fescue (F. campestris) grasslands in southwestern Alberta, aspen fleabane did not emerge from seed bank samples collected in April but did emerge from a single sample collected in October. Researchers evaluated emergence from samples gathered by vacuuming soil surfaces in April and October for 2 years. Emergence of aspen fleabane was 0.9 seedling/ft² (10/m²) from material collected at a site with moderate cattle use. Aboveground abundance of aspen fleabane in the study area was not reported (Willms and Quinton 1995).
Burned sites may be particularly conducive to aspen fleabane colonization. Abella (2009) found emergence was significantly greater when seeds were watered with a 10% aqueous smoke solution than with water alone (P < 0.05). Emergence was 27% from smoke-treated and 8% from untreated soil. Emergence was monitored for 60 days in a greenhouse kept at 75 ºF (24 ºC) with 14 hours of light. All seed had been stored at 23 ºF (-5 ºC) for 4 months following harvest (Abella 2009). In northern New Mexico, a single aspen fleabane seedling emerged from soils collected one year following a May fire in ponderosa pine (Pinus ponderosa) forests (Hunter and Omi 2006). It emerged from soils collected on low-severity burned areas. There was no emergence from collections made on moderately or severely burned areas, which were artificially seeded with cultivated grasses (Hunter and Omi 2006).
Impacts of 2004 pre-fire fuel reduction via thinning on vegetation recovery were examined 5 years following the 2011 Wallow fire in dry, mixed-conifer forest ecosystems in the Apache-Sitgreaves National Forest in eastern Arizona (Springer et al. 2018). Forests were primarily ponderosa pine, Douglas fir, and Gambel oak. Aspen fleabane was not classified as an indicator species on treated or untreated study areas at elevations from 7,927 to 9,311 ft (2,469 to 2,838 m) prior to the fire or in 2012. By 2016 composition of the treated and untreated areas were beginning to converge. Aspen fleabane’s presence as one of the indicator species in 2016 was attributed to seed dispersion by wind from offsite areas and more favorable conditions for forbs adapted to the increased insolation and temperatures experienced in the nontreated area (Springer et al. 2018).
Disturbance Ecology
Aspen fleabane abundance can be increased by fire but is often reduced by grazing (Moir 1967; Klemmedson 1956; Strahan et al. 2015). Frequency of aspen fleabane increased following fire and fire and thinning in ponderosa pine forests in east-central Arizona. Frequency of aspen fleabane averaged 0.7% before treatments and 5.3% five years after treatments on burned sites and 1% before and 7% five years after treatments on thinned and burned sites. Increases in aspen fleabane frequency also occurred on untreated sites but differences between the pre- and post-treatment years were smaller than those on treated sites (Strahan et al. 2015). Aspen fleabane was common (88% constancy, 5.5% cover) in spruce-fir forests in the Crested Butte area of west-central Colorado that burned 55 to 65 years earlier. Post-fire communities were dominated by blueberries (Vaccinium spp.) and Thurber’s fescue (Langenheim 1962). Aspen fleabane abundance was greatest on sites with moderate amounts of litter and bare soil when unburned, moderately burned, and severely burned ponderosa pine forests were compared in northern Arizona. Aspen fleabane was absent from sites with the lowest and highest amounts of litter and bare soil (Crawford et al. 2001).
Mattson et al. (2019) compared 20 stands that were clearcut 1 to 2 years prior to sampling with 32 unlogged controls in Engelmann spruce stands infested by spruce-beetle (Dendroctonus rufipennis) in the La Garita Mountains of southwestern Colorado. Aspen fleabane frequency was 34.4% and cover was 3.9% in the control (unlogged) areas. In the clearcuts, its frequency was 50% and cover was 3.3%.
Several studies found decreased abundance of aspen fleabane with grazing (Klemmedson 1956; Moir 1967; Eastmond 1968). Abundance of aspen fleabane was reduced by with either cattle or sheep grazing in subalpine Thurber’s fescue grasslands in southern New Mexico (Moir 1967). In Letterman’s needlegrass-Kentucky bluegrass (Poa pratensis) vegetation southeast of Cedar City, Utah, aspen fleabane was absent from sites with prolonged sheep grazing but production averaged 23 lbs/ac (26 kg/ha) on relatively ungrazed sites (Bowns and Bagley 1986). Aspen fleabane frequency increased from 26% to 54% after protection from grazing for 18 years in mountain brush (bigtooth maple-Gambel oak) vegetation at about 6,500 ft (1,980 m) elevation in Utah’s central Wasatch Mountains (Eastmond 1968). In subalpine Thurber’s fescue grasslands on the White River National Forest in western Colorado, aspen fleabane made up 1.2% of the vegetation composition on good, 2.8% on fair, and 0.9% on poor range condition plots (Klemmedson 1956).
Wildlife And Livestock Use
Aspen fleabane is eaten by mule deer (Odocoileus hemionus) (Deschamp et al. 1979), mountain goats (Oreamos americanus) (Chadwick 1974), northern pocket gophers (Thomomys talpoides) (Ward and Keith 1962), golden-mantled ground squirrels (Spermophilus lateralis) (Carleton 1966), and domestic sheep (Ellison 1954). It is also likely consumed and preferred by greater sage-grouse (Centrocercus urophasianus) (Dumroese et al. 2016).
Large mammals feed on aspen fleabane in summer. Aspen fleabane made up 18% of tame mule deer diets at a wet meadow site on the Ashley National Forest in northeastern Utah (Deschamp et al. 1979). Mountain goats fed on aspen fleabane in July, August, and September in subalpine vegetation in the Swan Mountains of western Montana (Chadwick 1974). Feeding observations were made from March 1971 to October 1972. Use was greatest in July when there were a total of 116 bites on aspen fleabane, which represented just 0.5% of total bites counted in July (Chadwick 1974).
Aspen fleabane is an important northern pocket gopher food and is also eaten, although not preferred, by golden-mantled ground squirrels. Aspen fleabane was the most important food for northern pocket gophers in grasslands in Black Mesa, west-central Colorado (Ward and Keith 1962). It was 24% by volume and 56% by occurrence in northern pocket gopher stomachs where aspen fleabane production averaged 72 lbs/ac (81 kg/ha) and it made up 5.2% of the vegetation composition. Northern pocket gophers consumed roots and aboveground plant parts. Consumption was greatest in July (Ward and Keith 1962). In captive feeding trials, northern pocket gophers consumed an average of 0.12 lb (53 g) of aspen fleabane daily (Tietjen et al. 1967). Consumption was even greater for herbicide (2,4-D)-treated plants (0.13 lb [57 g]/d). Herbicide treatments were targeting forbs in the area. Whole plants were collected from the mountain meadow on southwestern Colorado’s Black Mesa and fed to three northern pocket gophers from July 14 through August 11 (Tietjen et al. 1967). At rocky meadow sites near Gothic, Colorado, plants were sometimes eaten but not highly preferred by golden-mantled ground squirrels based on food choice tests and observations from June through September (Carleton 1966).
Aspen fleabane is highly palatable and preferred by domestic sheep. Its abundance is typically much less on grazed than ungrazed sites (Ellison 1954). Plants were absent from grassland sites subjected to prolonged sheep grazing but averaged 23 lbs/ac (26 kg/ha) production in relatively ungrazed grasslands near Cedar City, Utah (Bowns and Bagley 1986). On Utah’s Wasatch Plateau, aspen fleabane production was 0 lb/ac on sheep grazed, 5 lbs/ac (6 kg/ha) on plots protected from sheep but not from northern pocket gophers, and 39 lbs/ac (44 kg/ha) on plots protected from both grazers. Sites had been protected for about 30 years (Laycock and Richardson 1975).
Nutritional Value
For aspen fleabane seeds, oil content averaged 22% and protein content averaged 26.1% (Barclay and Earle 1974). Summer nutritional content of aspen fleabane growing in Idaho is reported for the summer months in Table 1 (Elliott and Flinders 1984).
Table 1. Average monthly nutrient and moisture content (%) of aspen fleabane growing in a cold meadow summer range in the River-of-No-Return Wilderness, Idaho (Elliott and Flinders 1984).
|
Month |
Crude fiber |
Crude protein |
Calcium |
Phosphorus |
Moisture |
|
——————————-%———————————– |
|||||
|
June |
18 |
19 |
0.56 |
0.16 |
75 |
|
July |
20 |
16 |
0.65 |
0.16 |
69 |
|
August |
24 |
13 |
1.03 |
0.20 |
58 |
Ethnobotany
Navajo and Thompson peoples may have used aspen fleabane medicinally. A decoction of aspen fleabane was used by the Navajo to ease menstrual pain and as a contraceptive (Moerman 2003). They may have also used it to treat head and abdominal pains (Shemluck 1982). The Thompson may have used fleabanes (Erigeron spp.) to treat sore throats and as a salve for other pain and swellings (Shemluck 1982).
Current Medicinal Use
The steam distilled fraction of aerial portions of aspen fleabane demonstrated antifungal and antimolluscicidal activity. This included the presence of phytochemicals toxic to the ram’s horn snail (Planobdela trivolvis). The snail is the intermediate host for a trematode, Bolbophorus confuses, that causes severe infections in pond-raised catfish in the Mississippi Delta (Meepagala et al. 2002). Potential value of the phytochemicals for control of the trematode is limited by their reported chemical instability.
Horticulture
Aspen fleabane is an attractive, pollinator friendly (Ley et al. 2007; Byrne et al. 2013), and easy to grow species. At least two cultivars are available commercially for ornamental use (Cornell University 2006; UW Ext. 2018). Many plant characteristics make aspen fleabane a desirable garden plant: it grows rapidly, tolerates most soils, has low to moderate water needs, grows well in both full sun and partial shade, is easy to propagate from seed or by division, has few known insect or disease problems, and it produces an abundance of attractive flowers (Mee et al. 2003; UW Ext. 2018).
Revegetation Use
Many aspen fleabane characteristics make it a good choice for restoration. It tolerates early seral conditions (Fig. 4) (Ellison 1949; Langenheim 1956), grows rapidly (Tilley et al. 2013), establishes with competition (S. Monsen, USFS retired, personal communication, July 2018), persists as a long-lived and somewhat shade-tolerant species (Louda et al. 1987; Caradonna and Bain 2016), occupies a wide range, and is adapted to a range of soil types and moisture regimes (see Soils section). The species attracts a diversity of pollinators (Watt et al. 1974; Wilson et al. 2010; Tilley et al. 2013), and its production of rhizomes and a taproot make it useful for soil stabilization (Shaw and Monsen 1983).

Figure 4. Aspen fleabane growing along a roadside in central Idaho. Photo: USFS.
Aspen fleabane is a potentially useful pollinator species for use in conservation, habitat improvement, and restoration in semideserts, open woodlands, coniferous forests, or alpine meadows in the mountains of Arizona and New Mexico (Ley et al. 2007). It provides habitat for monarch butterflies (Danaus plexippus) in the Southwest and is recommended for use on utility rights of way (Byrne et al. 2013).
Aspen fleabane has potential for use in mine reclamation (Brown et al. 2007). It was a volunteer species on phytotoxic, acidic, high-metal mine tailings in Leadville, Colorado, after sites were treated with biosolids and woody debris to attain target carbon:nitrogen ratios of 8:1 to 50:1. Mine waste was devoid of vegetation for 70 years. Treatments were applied in 2000, and aspen fleabane occurred on amended mine sites by 2005 although it was not seeded (Brown et al. 2007).
Developing A Seed Supply
For restoration to be successful, the right seed needs to be planted in the right place at the right time. Coordinated planning and cooperation is required among partners to first select appropriate species and seed sources and then properly collect, grow, certify, clean, store, and distribute seed for restoration (PCA 2015).
Developing a seed supply begins with seed collection from native stands. Collection sites are determined by current or projected revegetation requirements and goals. Production of nursery stock requires less seed than large-scale seeding operations, which may require establishment of agricultural seed production fields. Regardless of the size and complexity of any revegetation effort, seed certification is essential for tracking seed origin from collection through use (UCIA 2015).
Barga et al. (2020) analyzed collections of aspen fleabane and other priority forb species harvested by the BLM Seeds of Success program. Collected seeds are used for seed banking, immediate restoration, or seed increase in agricultural seed fields. The range of aspen fleabane was estimated to be 419,449 square miles (1,086,369 km2) with 7.1% impacted by fire from 2000 to 2017. Climate at the collection sites was compared to climate across the species range and to restoration sites within the species range. For the 44 aspen fleabane collections, average annual precipitation was greater at the collection sites than at the ecological sites, average summer precipitation and maximum temperature were lower, and average minimum temperature was similar. When collection sites were compared to the restoration sites, average annual and summer precipitation were greater at the collection sites, while average minimum and maximum temperatures were lower. Barga et al. (2020) emphasized the importance of carefully selecting populations for seed collection to provide supplies of adapted seed for disturbed areas requiring revegetation.
Seed Sourcing
Because empirical seed zones are not currently available for aspen fleabane, generalized provisional seed zones developed by Bower et al. (2014), may be used to select and deploy seed sources. These provisional seed zones identify areas of climatic similarity with comparable winter minimum temperature and aridity (annual heat:moisture index). In Figure 5, Omernik Level III Ecoregions (Omernik 1987) overlay the provisional seeds zones to identify climatically similar but ecologically different areas. For site-specific disturbance regimes and restoration objectives, seed collection locations within a seed zone and ecoregion may be further limited by elevation, soil type, or other factors.
The Western Wildland Environmental Threat Assessment Center’s (USFS WWETAC 2024) Threat and Resource Mapping (TRM) Seed Zone application provides links to interactive mapping features useful for seed collection and deployment planning. The Climate Smart Restoration Selection Tool (Richardson et al. 2020) can also guide restoration planning, seed collection, and seed deployment, particularly when addressing climate change considerations.
Occurrence Map
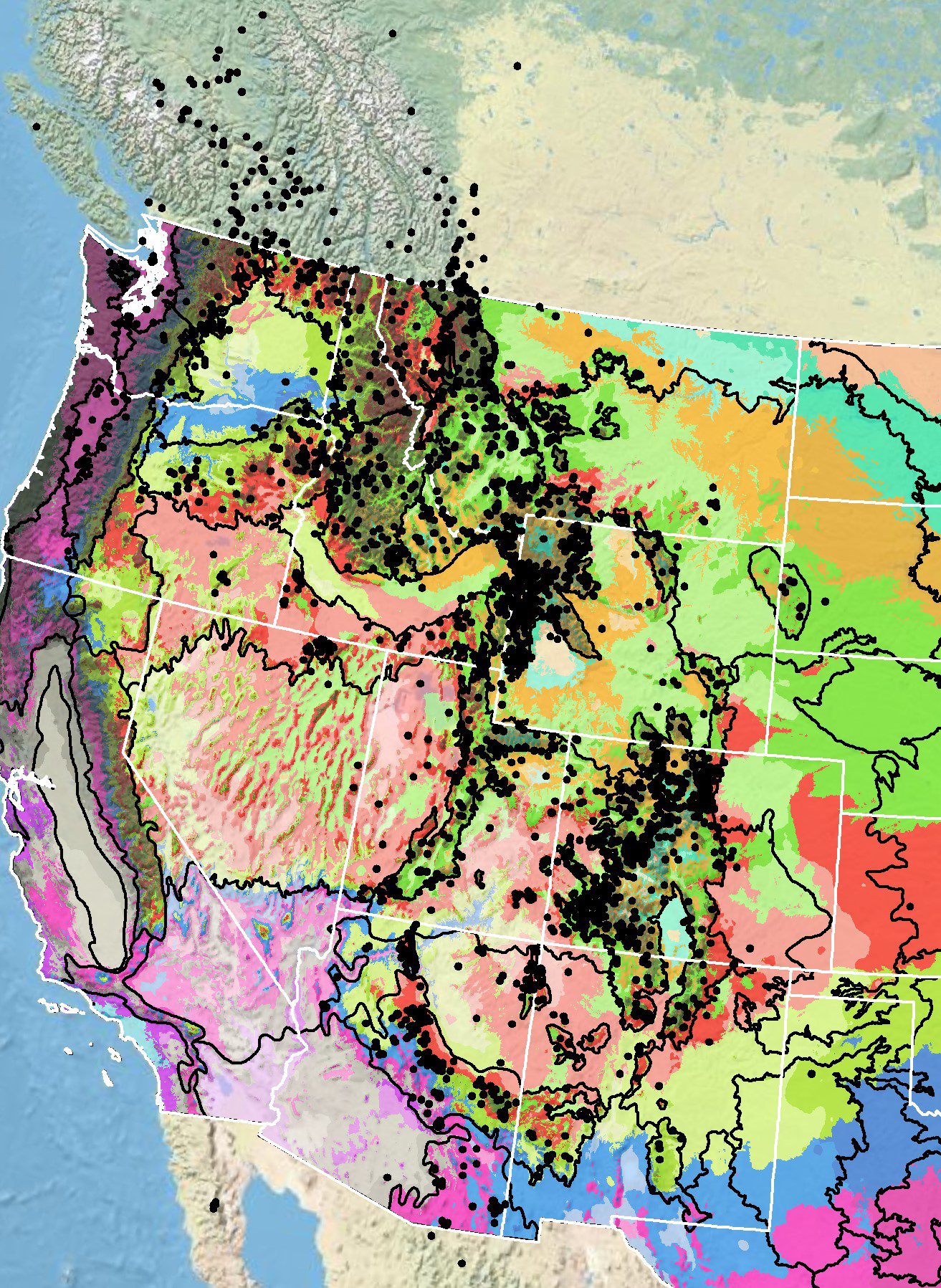
Figure 5. Distribution of aspen fleabane (black circles) based on geo-referenced herbarium specimens and observational data from 1870-2016 (CPNWH 2017; SEINet 2017; USGS 2017). Generalized provisional seed zones (colored regions; Bower et al. 2014) are overlain by Omernik Level III Ecoregions (black outlines) (Omernik 1987; USDI EPA 2017). Interactive maps, legends, and a mobile app are available (USFS WWETAC 2024). Map prepared by M. Fisk, USGS.
Releases
As of early 2025, there were no aspen fleabane germplasm releases for wildland restoration.
Wildland Seed Collection
Aspen fleabane seed can be harvested about a month after flowering, which occurs from July to September depending on location (Tilley et al. 2013, 2019). Seeds mature earlier at low-elevation than high-elevation sites (USDI BLM SOS 2017; S. Jackman, Eastern Oregon Stewardship Services [EOSS], personal communication, March 2018). Seed heads typically contain about 100 seeds that ripen somewhat uniformly (Mahalovich 2016). It is common, however, for mature seed heads to occur near the top of the plant while there are still flowers at the base. Seed maturity can also vary among plants within a population (Mahalovich 2016).
The harvest dates reported by the U.S. Department of the Interior, Bureau of Land Management’s Seeds of Success collection crews were July 1 to September 25 for 21 total collections made over 11 years (2002-2024) from elevations of 1,700 to 9,570 ft (520–2,920 m) at sites in Oregon, Washington, Montana, Idaho, Colorado, Utah, and Nevada. Most harvests (14 of 21) were made in August (USDI BLM SOS 2017, 2025). In a single year of harvests made by Eastern Oregon Stewardship Services, earliest flowers were found on June 30 at 4,200 ft (1,280 m) elevation and latest flowers on August 3 at 7,300 ft (2,230 m) elevation (S. Jackman, EOSS, personal communication, March 2018). Seed was collected on August 2 from the low-elevation and September 7 at the high-elevation site. Seed has also been found on completely cured plants, suggesting a long collection window (S. Jackman, EOSS, personal communication, March 2018).
In Oregon, aspen fleabane seed collections were made in forest openings, riparian areas, and along roadsides (Fig. 4), but plants were especially common and abundant on burned sites. On one burned site, aspen fleabane occupied several acres. Damage to aspen fleabane seed by fungal and insect pests was rare (S. Jackman, EOSS, personal communication, March 2018).
Several potentially co-occurring species are easily mistaken for aspen fleabane, especially at the seed harvesting stage (Fig. 6). Some of these are: subalpine fleabane (Erigeron peregrinus), threenerve fleabane, Unita fleabane (E. uintahensis), and western mountain aster (Symphyotrichum spathulatum). It is a good idea to have vouchers verified prior to collecting large quantities of seed (J. Irwin, USFS, personal communication, March 2018).
Wildland Seed Certification
Verification of species and tracking of geographic source is important whether wildland seed is collected for immediate project use or for cultivated increase. There is an official Source Identification process available for commercially produced seed that follows procedures established by the Association of Official Seed Certifying Agencies (AOSCA) Pre-Variety Germplasm Program (UCIA 2015; Young et al. 2020). Certification is optional but provides assurance of species identification and seed source for buyers. Seed certification is not always necessary, especially when seed is being locally produced and redistributed between cooperating agencies or parties. If wildland-collected seed is to be certified prior to sale for direct use in ecological restoration projects, collectors or those who contract for collection must apply for Source-Identified certification prior to making collections. Pre-collection applications, site inspections, and species and seed amount verification are handled by the AOSCA member state agency where seed collections will be made (see listings at AOSCA.org). Seed certification can increase seed price and marketability. It is important to become acquainted with state certification agency procedures, regulations, and deadlines.
If wildland collected seed is to be used to produce source-identified certified seed in seed production fields or nursery stock propagation (See Agricultural Seed Certification section), detailed information regarding collection site and collecting procedures are required. Wildland seed collections acquired within established protocols of recognized public agencies, however, are normally eligible to enter the certification process as stock seed without routine certification agency site inspections. Photos and herbarium specimens may be required. If seed is not certified by the collector or the contractor, this collection site information must be provided to the grower to enable certification.
Collection Timing

Figure 6. Ripening seed heads on aspen fleabane plants in Oregon. Photo: USDI BLM OR932 SOS.
Seeds are mature and ready for collection when they begin to turn brown and leak no moisture when squeezed (Mahalovich 2016). Because seeds are non-dormant and germinate readily with moisture, wet seed heads should be avoided and collections should only be made when seed heads are dry and humidity levels are low (See the Phenology section).
Collection Methods
Clipping mature seed heads into paper bags or other containers is recommended. Because aspen fleabane seed is available commercially, revegetated areas such as roadsides, mine sites, recreation areas, etc. should generally be avoided for wildland seed collecting (Mahalovich 2016). Although such areas may be suitable for collection if the seeding is well established and provenance of the seeded material is known and appropriate.
Several collection guidelines and methods should be followed to maximize the genetic diversity of wildland collections: 1) collect seed from a minimum of 50 randomly selected plants; 2) collect from widely separated individuals throughout a population without favoring the most robust or avoiding small statured plants; and 3) collect from all microsites including habitat edges (Basey et al. 2015). General collecting recommendations and guidelines are provided in online manuals (e.g., ENSCONET 2009; USDI BLM SOS 2023).
It is critical that wildland seed collection does not impact the sustainability of native plant populations. Collectors should take no more than 20% of the viable seed available at the time of harvest (USDI BLM SOS 2023). Additionally, care must be taken to avoid the inadvertent collection of weedy species, particularly those that produce seeds similar in shape and size to those of aspen fleabane.
Post-Collection Management
Thorough drying of aspen fleabane seed is critical for successful cleaning and storage (Fig. 7) (Mahalovich 2016). Seed molds if not dried carefully and quickly after harvest. Seed should be dried in an open area with good airflow before transporting and cleaning. Insect larvae present in the seed heads at the time of collection can impact seed yields before and after harvest and cleaning. If seed fill checks at the time of harvest suggest insect damage, freezing or pesticide treatments can be used to reduce seed yield losses (Mahalovich 2016).
Seed Cleaning
The tiny aspen fleabane seeds can be difficult to clean to high purity (Shaw and Monsen 1983). A hammermill and an air column separator are recommended for cleaning large seed lots, and small seed lots can be cleaned by rubbing the seeds over a screen to separate the seeds from their pappi (Fig. 8; Luna 2005).

Figure 7. Aspen fleabane seed (scale, mm). Photo: Sustainability in Prisons Project.

Figure 8. Cleaned aspen fleabane seed, most pappi removed. Photo: Sustainability in Prisons Project.
Seed lots collected by the Great Basin Native Plant Project, were cleaned by processing seed 1) through a small brush machine (mantle #32, speed 1, medium bristle brushes), 2) through a sieve or clipper, and 3) through an air column (J. Irwin, USFS, personal communication, March 2018).
Seed Storage
Dry seed (5-8% moisture) stored at temperatures of 32 to 36 °F (0-2 °C) will remain viable for many years (Huang 2015).
Seed Testing
There is no official AOSA procedure for examining viability, so viability of aspen fleabane is tested using the general procedures described for other Asteraceae genera. There is a rule for testing germination of aspen fleabane seed (AOSA 2016). Germination is tested at 59 °F (15 °C) with light. The first emergence count is made at 6 days and the last count at 16 days.
Purity Testing
Purity is evaluated using standard AOSA procedures. Minimum seed weight for purity testing is 0.8 g and for noxious-weed seed or bulk examination is 8 g. Approximately 3,010 seeds weigh 1 gram (AOSA 2016).
Germination Biology
Many studies report high germination of aspen fleabane seed over a range of temperatures with and without prechilling and regardless of light conditions (Springer 2001; Huang 2015; Kramer 2016). Although seed will germinate without pretreatment, cold stratification for 30 to 60 days encouraged more uniform germination of seed collected in Glacier National Park (Luna 2005). These different germination requirements suggest that germination conditions may vary by population or be affected by storage conditions and durations.
Laboratory testing (SER, INSR, RBGK 2024) obtained 100% germination after 7 days for seeds incubated on a 1% agar medium, exposed to an 8/16 hour light-dark cycle and a constant temperature of 59 °F (15 °C). Germination was slower (24 days) when the temperature was warmer (68 °F [20 °C]) and about 50% lower when the temperature was cooler (50 °F [10 °C]) (SER, INSR, RBGK 2024). In other studies on field-collected seed, 74% germination was achieved after 7 days and 98% germination after 21 days at alternating temperatures of 68/59 °F (20/15 °C) and a 8/16 hour light-dark cycle (Paulsen 1970). At the Rocky Mountain National Park Native Plant Nursery, aspen fleabane seed germinated uniformly, rapidly, and to high percentages at warm temperatures (55–70 °F [13–21 °C]) (Springer 2001; Huang 2015). Deno in Love et al. (2019) reported that germination occurred in 1 to 4 weeks for seeds incubated at 70 oF (21 oC).
Kramer (2016) found cooler temperatures resulted in greater germination than warmer temperatures for seed collected from the Colorado Plateau. Germination was nearly 100% when seed was incubated in a growth chamber set to reflect early spring conditions (4 wks at 52/34 °F [11/1 °C], 12 hr light/dark cycles). Germination was also high in simulated winter conditions (12 wks at 34 °F [1 °C] followed by 4 wks at 52–68 °F/34–50 °F [11–20 °C/1–10°C]). Germination was less than 50% after 4 weeks at 59/41 °F (15/5 °C) and less than 10% after 4 weeks at 68/50 °F (20/10 °C) (Kramer 2016).
In other studies, stratification impacted light requirements for germination (Hoffman 1985). Germination was significantly better when non-stratified seed was exposed to light (68%) than when kept in the dark (16%) at a constant temperature of 75 °F (24 °C) (P < 0.05). Seeds were collected from aspen woodlands in Routt County, Colorado, in late August or early September. For seeds given a long-duration stratification (120 d), germination was significantly greater in the dark (68%) than in the light (56%) (P < 0.05) (Hoffman 1985).
Wildland Seed Yield And Quality
Aspen fleabane seeds are tiny, ranging from about 1 to 3 million seeds/lb (2.2-6.6 million/kg) (Hassell et al. 1998; SER, INSR, RBGK 2024). Post-cleaning seed yield and quality of seed lots collected in the Intermountain region are provided in Table 2 (USFS BSE 2017, 2025). The results indicate that aspen fleabane seed can generally be cleaned to high levels of purity and that seed fill and viability of fresh seed is generally high.
Table 2. Seed yield and quality of aspen fleabane seed lots collected in the Intermountain region, cleaned by the Bend Seed Extractory, and tested by the Oregon State Seed Laboratory or the USFS National Seed Laboratory (USFS BSE 2017, 2025).
|
Characteristic |
Mean |
Range |
Samples (#) |
|
Bulk weight (lbs) |
0.86 |
0.06-5.13 |
23 |
|
Clean weight (lbs) |
0.14 |
0.005-1.46 |
23 |
|
Clean-out ratio |
0.14 |
0.03-0.32 |
23 |
|
Purity (%) |
95 |
75-99 |
23 |
|
Fill (%)¹ |
94 |
80-99 |
23 |
|
Viability (%)² |
94 |
79-98 |
17 |
|
Seeds/lb |
2,451,669 |
1,064,788-3,600,000 |
23 |
|
Pure live seeds/lb |
2,249,218 |
1,439,775-3,080,160 |
17 |
¹100 seed X-ray test
²Tetrazolium chloride test
Marketing Standards
Acceptable seed purity, viability, and germination specifications vary with revegetation plans. Purity needs are highest for precision seeding equipment like that used in nurseries, while some rangeland seeding equipment handles lower purity seed. Seed companies in Utah that buy, clean, and market wildland seed reported that they clean aspen fleabane seed to 80% purity (S. Monsen, USFS retired, personal communication, July 2018).
Agricultural Seed Production
Aspen fleabane produces seed in its first year under cultivation (Hottes cited in Bender et al. 2000) and is long-lived (Caradonna and Bain 2016), suggesting seed could possibly be harvested for several years, provided certification requirements are met. Detailed recommendations are lacking for establishment and care of aspen fleabane seed production fields. In 2018, wildland seed distributors located in Utah reported limited to no availability of wildland seed collections suggesting that seed production fields may be needed for this species (S. Monsen, USFS retired, personal communication, July 2018).
Field experiments suggest that fertilization would not benefit aspen fleabane stands and that certain herbicides could be used to control weeds in seed fields. In a Thurber’s fescue grassland, aspen fleabane decreased in abundance with additions of nitrogen and phosphorous (Paulsen 1970). In meadows near RMBL, nitrogen additions had no strong effect on per-flower reproduction (Burkle and Irwin 2010). In an evaluation of spotted knapweed (Centaurea stoebe subsp. micranthos) control and native forb recovery with herbicide only and fire followed by herbicide treatments, aspen fleabane density was not significantly affected by the treatments (P < 0.1) (Carpenter 1986). Study sites were invaded grasslands near Missoula, Montana. Herbicides tested were picloram, clopyralid, picloram plus clopyralid, and metsulfluron methyl. Although aspen fleabane density differences were not significant, there were observed patterns 13 months following treatments. More aspen fleabane plants occurred on burned and herbicide-treated than on unburned and herbicide-treated plots, and aspen fleabane appeared more sensitive to clopyralid and metsulfuron methyl than picloram (Carpenter 1986).
Aspen fleabane is a fungal host for Crocicreas nigrofuscum, Entyloma compositarum, Erysiphe cichoracearum, Puccinia dioicae, P. extensicola var. erigerontis, and Ramularia macrospora (Farr and Rossman 2025). Fleabanes are also noted hosts for gall-forming fungi (Asteromyia modesta) (Stireman et al. 2010). The extent to which these fungi may be problematic in seed field or nursery production was not reported. Any weed or fungal control measures in seed fields should be timed properly to avoid injury to the myriad of pollinators attracted to aspen fleabane flowers (see Pollination section).
Agricultural Seed Certification
In order to minimize genetic changes in specific accessions of native species when increased in cultivated fields, it is essential to track the geographic source and prevent inadvertent hybridization or selection pressure. This is accomplished by following third party seed certification protocols for Pre-Variety Germplasm (PVG) as established by the Association of Official Seed Certification Agencies (AOSCA). AOSCA members in the U.S., Canada, and other countries administer PVG requirements and standards that track the source and generation of planting stock. Field and cleaning facility inspections then monitor stand establishment, proper isolation distances, control of prohibited weeds, seed harvesting, cleaning, sampling, testing, and labeling for commercial sales (UCIA 2015; Young et al. 2020).
To produce certified seed, seed growers apply for certification of their production fields prior to planting and plant only certified stock seed of an allowed generation (usually less than four). The systematic and sequential tracking through the certification process requires preplanning, knowing state regulations and deadlines, and is most smoothly navigated by working closely with state certification agency personnel. See the Wildland Seed Certification section for more information on stock seed sourcing.
Nursery Practice
Aspen fleabane nursery stock has been grown successfully by the Glacier and Rocky Mountain National Park (GNP and RMNP) Plant Nurseries. At the GNP Nursery, firm root plugs with multiple true leaves were produced from cleaned aspen fleabane seed (Luna 2005). Seeds were sown into a nursery soil mix with timed release fertilizer and kept in a fully automated greenhouse. Containers were spaced to maintain good airflow. An emphasis was placed on keeping containers evenly moist during germination and establishment. Plants took 1 month to establish, the active growth phase was 2 months, and by mid- to late summer, seedlings were moved outdoors to harden (Springer 2001; Luna 2005). The hardening phase lasted 3 months, after which time plugs were either fall planted or overwintered with protective Microfoam sheets and outplanted in the spring (Luna 2005).
At the RMNP Nursery in Estes Park, Colorado, seedlings were grown from seed collected at McGraw Ranch (Huang 2015). Seed was manually sown in a 4 × 8 flats and covered lightly with a superfine soil mix. Seed germinated uniformly after 7 days in a greenhouse at 65 to 70 °F (18-21 °C) during the day and 55 °F (13 °C) at night. Germination containers were kept under tents on heated pads (70 °F [21 °C]). Containers were misted for about 15 minutes every 12 hours. After seeds germinated, the tents were removed but misting continued for another week. After this, seedlings were watered deeply every other day and provided water-soluble fertilizer with micro-nutrients each week for a month. Plants were moved to a cold frame in late March or early April to harden for 1 to 3 weeks prior to planting (Huang 2015).
Wildland Seeding And Planting
Aspen fleabane can be broadcast seeded if seed is subsequently covered. It can also be drill seeded or cultipacker seeded as part of a mixture (Shaw and Monsen 1983). Field observations and greenhouse experiments suggest aspen fleabane may do well in seeding mixtures (Ellison 1949). In surveys of denuded and patchy vegetation recovery on the Wasatch Plateau, central Utah, aspen fleabane was frequently found establishing where penstemon or yarrow plants had already established. Once established, aspen fleabane spread out beyond the penstemon and yarrow patches (Ellison 1949).
In greenhouse experiments, aspen fleabane survival was high when present at a very low level in a seed mixture (Paulsen 1970). Five-gallon containers were seeded with 192 total seeds and varying proportions of 2 grass species (Thurber’s fescue and Letterman’s needlegrass, 44 to 86% of seeds) and 3 forb species (Fremont’s geranium [Geranium caespitosum var. fremontii], beautiful cinquefoil [Potentilla pulcherrima], and aspen fleabane, 14-56% of seeds). The total number of aspen fleabane seeds planted per container ranged from 4 to 16. Survival of aspen fleabane ranged from 87 to 100% after 267 days in normal water treatments. Survival was lower (57-97%) in low water treatments and when just four aspen fleabane seeds were present in the containers (Paulsen 1970).
In restoration of several disturbed sites in the Manti-La Sal and Uinta-Wasatch-Cache National Forests of Utah, aspen fleabane was successfully seeded using an imprint seeder (S. Monsen, USFS retired, personal communication, July 2018). Seeding with conventional seeders, which planted seeds too deep (> 0.5 in), resulted in little to no establishment. Aspen fleabane was seeded with about 20 other tall forb species at the restoration sites and across all sites remained a dominant species for at least 5 years after seeding (S. Monsen, USFS retired, personal communication, July 2018).
Aspen fleabane survival was good from nursery stock (plugs) planted on the Rocky Prairie Preserve south of Olympia, Washington (Thomas and Gamon 1997). Survival of aspen fleabane outplanted in March remained high by late May of the same year. Monitoring in the following years revealed that plants established well, reached reproductive maturity, and produced new seedlings (Thomas and Gamon 1997).
Acknowledgements
Funding for the Western Forbs: Biology, Ecology, and Use in Restoration was provided by the USDI BLM Great Basin Native Plant Materials Ecoregional Program through the Great Basin Fire Science Exchange. We very much appreciate the review of this chapter by Steve Monsen USFS (retired) and Berta Youtie, Eastern Oregon Stewardship Services.
This research was supported in part by the USDA Forest Service, Rocky Mountain Research Station. The findings and conclusions in this publication are those of the authors and should not be construed to represent any official USDA or U.S. Government determination or policy.
Literature Cited
Abella, S.R. 2009. Smoke-cued emergence in plant species of ponderosa pine forests. Fire Ecology. 5(1): 22-37.
Allman, V.P. 1953. A preliminary study of the vegetation in an exclosure in the chaparral of the Wasatch Mountains, Utah. Utah Academy Proceedings. 30: 63-78.
Association of Official Seed Analysts [AOSA]. 2016. AOSA rules for testing seeds. Vol. 1. Principles and procedures. Washington, DC: Association of Official Seed Analysts.
Association of Official Seed Analysts [AOSA]. 2023. AOSA rules for testing seeds. Vol. 1. Principles and procedures. Washington, DC: Association of Official Seed Analysts.
Barclay, A.S.; Earle, F.R. 1974. Chemical analyses of seeds III: Oil and protein content of 1253 species. Economic Botany. 28(2): 178-236.
Barga, S.C.; Olwell, P.; Edwards, F.; Prescott, L.; Leger, E.A. 2020. Seeds of Success: A conservation and restoration investment in the future of U.S. lands. Conservation Science and Practice. 2(7): e209.
Basey, A.C.; Fant, J.B.; Kramer, A.T. 2015. Producing native plant materials for restoration: 10 rules to collect and maintain genetic diversity. Native Plants Journal. 16(1): 37-53.
Bender, M.H.; Baskin, J.M.; Baskin, C.C. 2000. Age of maturity and life span in herbaceous, polycarpic perennials. Botanical Review. 66(3): 311-349.
Boggs, C.L.; Inouye, D.W. 2012. A single climate driver has direct and indirect effects on insect population dynamics. Ecology Letters. 15(5): 502-508.
Bower, A.D.; St. Clair, J.B.; Erickson, V. 2014. Generalized provisional seed zones for native plants. Ecological Applications. 24(5): 913-919.
Bowers, J.E.; McLaughlin, S.P. 1987. Flora and vegetation of the Rincon Mountains, Pima County, Arizona. Desert Plants. 8(2): 50-94.
Bowns, J.E.; Bagley, C.F. 1986. Vegetation responses to long-term sheep grazing on mountain ranges. Journal of Range Management. 39(5): 431-434.
Braun, A.F. 1921. Two weeks collecting in Glacier National Park. Proceedings of the Academy of Natural Sciences of Philadelphia. 73(1): 1-23.
Brown, S.; DeVolder, P.; Compton, H.; Henry, C. 2007. Effect of amendment C:N ratio on plant richness, cover and metal content for acidic Pb and Zn mine tailings in Leadville, Colorado. Environmental Pollution. 149(2): 165-172.
Burkle, L.A.; Glenny, W.R.; Runyon, J.B. 2020b. Intraspecific and interspecific variation in floral volatiles over time. Plant Ecology. 221(7): 529-544.
Burkle, L.A.; Irwin, R.E. 2009. The importance of interannual variation and bottom-up nitrogen enrichment for plant-pollinator networks. Oikos. 118(12): 1816-1829.
Burkle, L.A.; Irwin, R.E. 2010. Beyond biomass: Measuring the effects of community-level nitrogen enrichment on floral traits, pollinator visitation and plant reproduction. Journal of Ecology. 98(3): 705-717.
Byrne, M.; Davies Adams, L.; Wojcik, V. 2013. Monarch habitat development on utility rights of way: Southwest. San Francisco, CA: Pollinator Partnership. 32 p.
Caradonna, P.J.; Bain, J.A. 2016. Frost sensitivity of leaves and flowers of subalpine plants is related to tissue type and phenology. Journal of Ecology. 104(1): 55-64.
Carleton, W.M. 1966. Food habits of two sympatric Colorado sciurids. Journal of Mammalogy. 47(1): 91-103.
Carpenter, J.L. 1986. Responses of three plant communities to herbicide spraying and burning of spotted knapweed (Centaurea maculosa) in western Montana. Missoula, MT: University of Montana. Thesis. 110 p.
Chadwick, D.H. 1974. Mountain goat ecology: Logging relationships in the Bunker Creek drainage of western Montana. Missoula, MT: University of Montana. Thesis. 262 p.
Consortium of Pacific Northwest Herbaria [CPNWH]. 2017. Seattle, WA: University of Washington Herbarium, Burke Museum of Natural History and Culture. pnwherbaria.org/
Cornell University. 2006. Flower growing guides – Erigeron speciosus. Ithaca, NY: Cornell University. http://www.gardening.cornell.edu/homegardening/scenedb99.html [Accessed 2018 January 3].
Costello, D.F. 1944. Important species of the major forage types in Colorado and Wyoming. Ecological Monographs. 14(1): 107-134.
Craighead, J.J.; Craighead, F.C. Jr.; Davis, R.J. 1963. A field guide to Rocky Mountain wildflowers from northern Arizona and New Mexico to British Columbia. Boston, MA: Houghton Mifflin Company. 277 p.
Crawford, J.A.; Wahren, C.H.A.; Kyle, S.; Moir, W.H. 2001. Responses of exotic plant species to fires in Pinus ponderosa forests in northern Arizona. Journal of Vegetation Science. 12(2): 261-268.
Cronquist, A. 1943. The North American species of Erigeron centering about E. speciosus (Lindl.) DC. and E. glabellus Nutt. Bulletin of the Torrey Botanical Club. 70(3): 265-274.
Cronquist, A.; Holmgren, A.H.; Holmgren, N.H.; Reveal, J.L.; Holmgren, P.K. 1994. Intermountain flora: Vascular plants of the Intermountain West, U.S.A. Volume 5: Asterales. Bronx, NY: The New York Botanic Garden. 496 p.
de Valpine, P.; Harte, J. 2001. Plant responses to experimental warming in a montane meadow. Ecology. 82(3): 637-648.
Deschamp, J.A.; Urness, P.J.; Austin, D.D. 1979. Summer diets of mule deer from lodgepole pine habitats. The Journal of Wildlife Management. 43(1): 154-161.
Discover Life. 2025. Sam Houston State University. Available: https://www.discoverlife.org/
Drake, D.; Ewing, K. nd. Germination requirements of 32 native Washington prairie species. Seattle, WA: The Nature Conservancy: 181-190.
Dumroese, R.K.; Luna, T.; Pinto, J.R.; Landis, T.D. 2016. Forbs: Foundation for restoration of monarch butterflies, other pollinators, and greater sage-grouse in the western United States. Native Plants Journal. 36(4): 499-511.
Dunne, J.A.; Harte, J.; Taylor, K.J. 2003. Subalpine meadow flowering phenology responses to climate change: Integrating experimental and gradient methods. Ecological Monographs. 73(1): 69-86.
Eastmond, R.J. 1968. Vegetational changes in a mountain brush community of Utah during eighteen years. Provo, UT: Brigham Young University. Thesis. 261 p.
Elliott, C.L.; Flinders, J.T. 1984. Plant nutrient levels on two summer ranges in the River of No Return Wilderness Area, Idaho, USA. The Great Basin Naturalist. 44(4): 621-626.
Ellison, L. 1949. Establishment of vegetation on depleted subalpine range as influenced by microenvironment. Ecological Monographs. 19(2): 95-121.
Ellison, L. 1954. Subalpine vegetation of the Wasatch Plateau, Utah. Ecological Monographs. 24(2): 89-184.
European Native Seed Conservation Network [ENSCONET]. 2009. ENSCONET seed collecting manual for wild species. Edition 1: 32 p.
Fall, P.L. 1992. Pollen accumulation in a montane region of Colorado, USA: A comparison of moss polsters, atmospheric traps, and natural basins. Review of Palaeobotany and Palynology. 72(3-4): 169-197.
Fall, P.L. 1997. Fire history and composition of the subalpine forest of western Colorado during the Holocene. Journal of Biogeography. 24(3): 309-325.
Farr, D.F.; Rossman, A.Y. 2025. Fungal databases, U.S. National Fungus Collections. U.S. Department of Agriculture, Agricultural Research Service. https://fungi.ars.usda.gov/
Franklin, J.F.; Dyrness, C.T. 1973. Natural vegetation of Oregon and Washington. Gen. Tech. Rep. PNW-8. Portland, OR: U.S. Department of Agriculture, Forest Service, Pacific Northwest Forest and Range Experiment Station. 452 p.
Glenny, W.; Runyon, J.; Burkle, L. 2022. Assessing pollinator friendliness of plants and designing mixes to restore habitat for bees. Gen Tech. Rep. RMRS-GTR-429. Fort Collins, CO: USDA Forest Service, Rocky Mountain Research Station. 52 p.
Harte, J. 2019. Reflections on 27 years of manipulated ecosystem warming in a subalpine meadow. In: Mohan, J.E., ed. Ecosystem consequences of soil warming. London, England: Academic Press: 1-27.
Hassell, W.; Beavers, W.R.; Ouellette, S.; Mitchell, T. 1998. Seeding rate statistics for native and introduced species. Plant Materials Technical Note 21. Portland, OR: U.S. Department of Agriculture, Natural Resources Conservation Service. 25 p.
Hermann, F. 1966. Notes on western range forbs: Cruciferae through Compositae. Agric. Handb. 293. Washington, DC: U.S. Department of Agriculture, Forest Service. 365 p.
Hersperger, A.M.; Forman, R.T.T. 2003. Adjacency arrangement effects on plant diversity and composition in woodland patches. Oikos. 101(2): 279-290.
Hoffman, G.R. 1985. Germination of herbaceous plants common to aspen forests of western Colorado. Bulletin of the Torrey Botanical Club. 112(4): 409-413.
Holiday, S. 2000. A floristic study of Tsegi Canyon, Arizona. Madroño. 47(1): 29-42.
Huang, J. 2015. Plant propagation protocol for Erigeron speciosus: ESRM 412 Native plant production. U.S. Department of Agriculture, Natural Resource Conservation Service. 4 p.
Hunter, M.E., Omi, P.N. 2006. Seed supply of native and cultivated grasses in pine forests of the southwestern United States and the potential for vegetation recovery following wildfire. Plant Ecology. 183(1): 1-8.
Ingold, D.D.; CaraDonna, P.J.; Dooley, J.L.; Ingold, D.J.; Iler, A.M. 2024. Reproductive ecology of Erigeron speciosus, a montane perennial herb: Evidence of self-incompatibility. The Southwestern Naturalist. 68(1): 13-19.
Inouye, D.W. 2008. Effects of climate change on phenology, frost damage, and floral abundance of montane wildflowers. Ecology. 89(2): 353-362.
Jones, A.G.; Young, D.A. 1983. Generic concepts of aster (Asteraceae): A comparison of cladistic, phenetic, and cytological approaches. Systematic Botany. 8(1): 71-84.
Klemmedson, J.O. 1956. Interrelations of vegetation, soils and range conditions induced by grazing. Journal of Range Management. 9(3): 134-138.
Kramer, A. 2016. Propagation protocol for production of propagules (seeds, cuttings, poles, etc.) Erigeron speciosus (Lindl.) DC. seeds. Native Plant Network. U.S. Department of Agriculture, Forest Service, National Center for Reforestation, Nurseries, and Genetic Resources. http://npn.rngr.net/propagation/protocols [Accessed 2018 January 3].
Lady Bird Johnson Wildflower Center [LBJWC]. 2018. Erigeron speciosus (Lindl.) DC. Native Plant Database. Austin, TX: Lady Bird Johnson Wildflower Center. https://www.wildflower.org/plants-main [Accessed 2018 January 3].
Lambrecht, S.C.; Loik, M.E.; Inouye, D.W.; Harte, J. 2007. Reproductive and physiological responses to simulated climate warming for four subalpine species. New Phytologist. 173(1): 121-134.
Langenheim, J.H. 1956. Plant succession on a subalpine earthflow in Colorado. Ecology. 37(2): 301-317.
Langenheim, J.H. 1962. Vegetation and environmental patterns in the Crested Butte Area, Gunnison County, Colorado. Ecological Monographs. 32(3): 249-285.
Laycock, W.A.; Richardson, B.Z. 1975. Long-term effects of pocket gopher control on vegetation and soils of a subalpine grassland. Journal of Range Management. 28(6): 458-462.
Lesica, P. 2012. Manual of Montana vascular plants. Fort Worth, TX: Botanical Research Institute of Texas (BRIT) Press. 779 p.
Ley, E.; Buchmann, S.; Stritch, L.; Soltz, G. 2007. Selecting plants for pollinators: A regional guide for farmers, land managers, and gardeners in the Intermountain Semidesert Province. San Francisco, CA: Pollinator Partnership. 24 p.
Loik, M.E.; Harte, J. 1997. Changes in water relations for leaves exposed to a climate-warming manipulation in the Rocky Mountains of Colorado. Environmental and Experimental Botany. 37: 115-123.
Louda, S.M.; Dixon, P.M.; Huntly, N. 1987. Herbivory in sun versus shade at a natural meadow-woodland ecotone in the Rocky Mountains USA. Vegetatio. 72(3): 141-149.
Love, S.L.; Akins, C.J. 2019. Second summary of the native seed germination studies of Norman C Deno: Species with names beginning with letters C through E. Native Plants Journal. 20(1): 65-97.
Luna, T. 2005. Propagation protocol for production of container (plug) Erigeron speciosus (Lindl.) DC. plants 116 ml (7.0 cu. in). Native Plant Network. U.S. Department of Agriculture, Forest Service, National Center for Reforestation, Nurseries, and Genetic Resources. http://npn.rngr.net/propagation/protocols [Accessed 2018 January 3].
Mahalovich, M. 2016. Showy fleabane (Erigeron speciosus var. macrantha [Nutt] Cronquist and var. speciosus [Lindl.] DC). USFS native plant material development for sage-grouse chicks. On file at: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station, Boise, ID. 8 p.
Mansfield, D.H. 2000. Flora of Steens Mountain. Corvallis, OR: Oregon State University Press. 410 p.
Mattson, L.R.; Coop, J.D.; Battaglia, M.A.; Cheng, A.S.; Sibold, J.S.; Viner, S. 2019. Post-spruce beetle timber salvage drives short-term surface fuel increases and understory vegetation shifts. Forest Ecology and Management. 437: 348-359.
Mee, W.; Barnes, J.; Kjelgren, R.; Sutton, R.; Cerny, T.; Johnson, C. 2003. Water wise: Native plants for Intermountain landscapes. Logan, UT: Utah State University Press. 220 p.
Meepagala, K.M.; Sturtz, G.; Wise, D.; Wedge, D.E. 2002. Molluscicidal and antifungal activity of Erigeron speciosus steam distillate. Pest Management Science. 58: 1043-1047.
Moerman, D. 2003. Native American ethnobotany: A database of foods, drugs, dyes, and fibers of Native American peoples, derived from plants. Dearborn, MI: University of Michigan. http://naeb.brit.org
Moir, W.H. 1967. The subalpine tall grass, Festuca thurberi, community of Sierra Blanca, New Mexico. The Southwestern Naturalist. 12(3): 321-328.
Moss, E.H. 1955. The vegetation of Alberta. Botanical Review. 21(9): 493-567.
Mueggler, W.F.; Bartos, D.L. 1977. Grindstone Flat and Big Flat exclosures: A 41-year record of changes in clearcut aspen communities. Res. Pap. INT-195. Ogden, UT: U.S. Department of Agriculture, Forest Service. Intermountain Forest and Range Experiment Station. 24 p.
Neely, E.E.; Barkworth, M.E. 1984. Vegetation on soils derived from dolomite and quartzite in the Bear River Range, Utah: A comparative study. Bulletin of the Torrey Botanical Club. 111(2): 179-192.
Nesom, G.L. 2006. 186. Erigeron. In: Flora of North America Editorial Committee, ed. Flora of North America North of Mexico. Volume 20 Magnoliophyta: Asteridae, part 7: Asteraceae, part 2 Asterales, part 2 (Aster order). New York, NY: Oxford University Press: 256-348.
Omernik, J.M. 1987. Ecoregions of the conterminous United States. Map (scale 1:7,500,000). Annals of the Association of American Geographers. 77(1): 118-125.
Pardee, G.L.; Inouye, D.W.; Irwin, R.E. 2018. Direct and indirect effects of episodic frost on plant growth and reproduction in subalpine wildflowers. Global Change Biology. 24(2): 848-857.
Paulsen, H.A. 1970. The ecological response of species in a Thurber fescue community to manipulative treatments. Fort Collins, CO: Colorado State University. Thesis. 145 p.
Pavek, P.; Erhardt, B.; Heekin, T.; Old, R. 2012. Forb seedling identification guide for the Inland Northwest: Native, introduced, invasive, and noxious species. Pullman, WA: U.S. Department of Agriculture, Natural Resources Conservation Service, Pullman Plant Materials Center. 144 p.
Plant Conservation Alliance [PCA]. 2015. National seed strategy for rehabilitation and restoration 2015-2020. Washington, DC: U.S. Department of the Interior, Bureau of Land Management. 52 p.
Pleasants, J.M. 1980. Competition for bumblebee pollinators in Rocky Mountain plant communities. Ecology. 61(6): 1446-1459.
Pleasants, J.M. 1981. Bumblebee response to variation in nectar availability. Ecology. 62(6): 1648-1661.
Pohl, N.B.; Van Wyk, J.; Campbell, D.R. 2011. Butterflies show flower colour preferences but not constancy in foraging at four plant species. Ecological Entomology. 36(3): 280-300.
Ranne, B.M.; Baker, W.L.; Andrews, T.; Ryan, M.G. 1997. Natural variability of vegetation, soils, and physiography in the bristlecone pine forests of the Rocky Mountains. The Great Basin Naturalist. 57(1): 21-37.
Reed, J.F. 1952. The vegetation of the Jackson Hole Wildlife Park, Wyoming. The American Midland Naturalist. 48(3): 700-729.
Reynolds, L.V.; Cooper, D.J. 2011. Ecosystem response to removal of exotic riparian shrubs and a transition to upland vegetation. Plant Ecology. 212(8): 1243-1261.
Richardson, B.; Kilkenny, F.; St. Clair, B.; Stevenson-Molnar, N. 2020. Climate Smart Restoration Tool. https://climaterestorationtool.org/csrt/
Richardson, B.A.; Massatti, R.; Islam-Faridi, N.; Johnson, S.; Kilkenny, F.F. 2022. Assessing population genomic structure and polyploidy: A crucial step for native plant restoration. Restoration Ecology. 31(3): e13729
Riser, J.P.; Meyers, S.C. 2020. Erigeron (Fleabane, daisy). In: Meyers, S.C.; Jaster, T.; Mitchell, K.E.; Harvey, T.; Hardison, L.K., eds. Flora of Oregon, Volume 2: Dicots A-F. Fort Worth, TX: Botanical Research Institute of Texas (BRIT) Press: 249-262.
Robberecht, R.; Caldwell, M.M. 1987. Leaf epidermal transmittance of ultraviolet radiation and its implications for plant sensitivity to ultraviolet-radiation induced injury. Oecologia. 32(3): 277-287.
Robinson, G.S.; Ackery, P.R.; Kitching, I.; Beccaloni, G.W.; Hernández, L.M. 2025. HOSTS: A Database of the World’s Lepidopteran Hostplants [Data set]. Natural History Museum. https://doi.org/10.5519/havt50xw
SEINet – Regional Networks of North American Herbaria Steering Committee [SEINet]. 2017. SEINet Regional Networks of North American Herbaria. https://symbiota.org/seinet
Shaw, N.; Monsen, S.B. 1983. Nonleguminous forbs for rangeland sites. In: Monsen, S.B; Shaw, N. Managing Intermountain rangelands-Improvement of range and wildlife habitats; 1981 September 15-17; Twin Falls, ID; 1982 June 22-24; Elko, NV. Gen. Tech. Rep. INT-157. Ogden, UT: U.S. Department of Agriculture, Forest Service, Intermountain Forest and Range Experiment Station: 123-131.
Shaw, R.J. 1995. Utah wildflowers: A field guide to northern and central mountains and valleys. Logan, UT: Utah State University Press. 218 p.
Shemluck, M. 1982. Medicinal and other uses of the Compositae by Indians in the United States and Canada. Journal of Ethnopharmacology. 5: 303-358.
Shiflet, T.N. 1994. Rangeland cover types of the United States. Denver, CO: Society for Range Management. 152 p.
Slominski, A.H.; German, Z.; Burkle, L.A. 2018. Growth and physiological responses of subalpine forbs to nitrogen and soil moisture. Plant Ecology. 219(8): 941-956.
Society for Ecological Restoration; International Network for Seed Based Restoration; Royal Botanic Gardens [SER, INSR, RBGK]. 2024. Seed Information Database (SID). Available from: https://ser-sid.org/
Spellenberg, R. 2001. National Audubon Society field guide to North American wildflowers: Western region, revised edition. New York, NY: Alfred A. Knopf, Inc. 862 p.
Springer, J.D.; Huffman, D.W.; Stoddard, M.T.; Sánchez Meador, A.J.; Waltz, A.E.M. 2018. Plant community dynamics following hazardous fuel treatments and mega-wildfire in a warm-dry mixed-conifer forest of the USA. Forest Ecology and Management. 429(2018): 278-286.
Springer, L. 2001. Propagation protocol for production of propagules (seeds, cuttings, poles, etc.) Erigeron speciosus (Lindl.) DC. seeds. Native Plant Network. U.S. Department of Agriculture, Forest Service, National Center for Reforestation, Nurseries, and Genetic Resources. http://npn.rngr.net/propagation/protocols [Accessed 2018 January 3].
Stireman III, J.O.; Devlin, H.; Carr, T.G; Abbot, P. 2010. Evolutionary diversification of the gall midge genus Asteromyia (Cecidomyiidae) in a multitrophic ecological context. Molecular Phylogenetics and Evolution. 54(1): 194-210.
Strahan, R.T.; Stoddard, M.T.; Springer, J.D.; Huffman, D.W. 2015. Increasing weight of evidence that thinning and burning treatments help restore understory plant communities in ponderosa pine forests. Forest Ecology and Management. 353(1): 208-220.
Thomas, T.; Gamon, J. 1997. Restoration of a prairie plant community: Help for a threatened species. In: Warwick, C., ed. Fifteenth North American Prairie Conference. Bend, OR: The Natural Areas Association: 244-248.
Tietjen, H.P.; Halvorson, C.H.; Hegdal, P.L.; Johnson, A.M. 1967. 2,4-D herbicide, vegetation, and pocket gopher relationships Black Mesa, Colorado. Ecology. 48(4): 634-643.
Tilley, D.; Spencer, J.; Cracroft, T.; Brazee, B.; Vaughan, M.; Cruz, J.K. 2019. Creation and management of Utah butterfly habitat. Tech. Note Plant Materials 73. Boise, ID: U.S. Department of Agriculture, Natural Resources Conservation Service. 20 p.
Tilley, D.; Taliga, C.; Burns, C.; St. John, L. 2013. Plant materials for pollinators and other beneficial insects in eastern Utah and western Colorado. Tech. Note 2C. Boise, ID: U.S. Department of Agriculture, Natural Resources Conservation Service. 54 p.
University of Wyoming Extension [UW Ext.] 2018. Native plants for the Intermountain West. Laramie, WY: University of Wyoming. http://www.wyoextension.org/westernnativeplants
USDA Forest Service [USDA FS] 1937. Range plant handbook. Washington, DC: U.S. Department of Agriculture, Forest Service. 816 p.
USDA Forest Service, Bend Seed Extractory [USDA FS BSE]. 2017. Nursery Management Information System Version 4.1.11. Local Source Report 34-Source Received. Bend, OR: U.S. Department of Agriculture, Forest Service, Bend Seed Extractory.
USDA Forest Service, Bend Seed Extractory [USDA FS BSE]. 2025. Nursery Management Information System Version 4.1.11. Local Source Report 34-Source Received. Bend, OR: U.S. Department of Agriculture, Forest Service Bend Seed Extractory.
USDA Forest Service, Western Wildland Environmental Threat Assessment Center [USFS WWETAC]. 2024. TRM Seed Zone Applications. Prineville, OR: U.S. Department of Agriculture, Forest Service, Western Wildland Environmental Threat Assessment Center. https://research.fs.usda.gov/pnw/products/dataandtools/seed-zone-webmap
USDA Natural Resources Conservation Service [USDA NRCS]. 2025. The PLANTS Database. Greensboro, NC: U.S. Department of Agriculture, Natural Resources Conservation Service, National Plant Data Team. https://plants.sc.egov.usda.gov/
USDI Bureau of Land Management, Seeds of Success [USDI BLM SOS]. 2017. Seeds of Success collection data. Washington, DC: U.S. Department of the Interior, Bureau of Land Management, Plant Conservation and Restoration Program.
USDI Bureau of Land Management, Seeds of Success [USDI BLM SOS]. 2023. Bureau of Land Management technical protocol for the collection, study, and conservation of seeds from native plant species for Seeds of Success. Washington, DC: U.S. Department of the Interior, Bureau of Land Management, Plant Conservation and Restoration Program. 45 p.
USDI Bureau of Land Management, Seeds of Success [USDI BLM SOS]. 2025. Seeds of Success collection data. Washington, DC: U.S. Department of the Interior, Bureau of Land Management, Plant Conservation and Restoration Program.
USDI Environmental Protection Agency [USDI EPA]. 2017. Ecoregions. Washington, DC: U.S. Department of the Interior, Environmental Protection Agency. https://www.epa.gov/eco-research/ecoregions
USDI Geological Survey [USDI USGS]. 2017. Biodiversity Information Serving Our Nation (BISON). U.S. Geological Survey. https://www.gbif.us/
Utah Crop Improvement Association [UCIA]. 2015. How to be a seed connoisseur. Logan, UT: UCIA, Utah Department of Agriculture and Food, Utah State University and Utah State Seed Laboratory. 16 p.
Wadgymar, S.M.; Ogilvie, J.E.; Inouye, D.W.; Weis, A.E.; Anderson, J.T. 2018. Phenological responses to multiple drivers under climate change: insights from a long-term observational study and a manipulative field experiment. New Phytologist. 218(2): 517-529.
Wahlberg, N. 2001. The phylogenetics and biochemistry of host-plant specialization in Melitaeine butterflies (Lepidoptera: Nymphalidae). Evolution. 55(3): 522-537.
Ward, A.L., Keith, J.O. 1962. Feeding habits of pocket gophers on mountain grasslands, Black Mesa, Colorado. Ecology. 43(4): 744-749.
Watt, W.B.; Hoch, P.C.; Mills, S.G. 1974. Nectar resource use by Colias butterflies. Chemical and visual aspects. Oecologia. 14(4): 353-374.
Weaver, T.; Collins, D. 1977. Possible effects of weather modification (increased snowpack) on Festuca idahoensis meadows. Journal of Range Management. 30(6): 451-456.
Weber, W.A. 1976. Rocky Mountain flora. Boulder, CO: Colorado Associated University Press. 479 p.
Welsh, S.L. 1983. Utah flora: Compositae (Asteraceae). Great Basin Naturalist. 43(2): 179-357.
Welsh, S.L.; Atwood, N.D.; Goodrich, S.; Higgins, L.C., eds. 1987. A Utah flora. The Great Basin Naturalist Memoir 9. Provo, UT: Brigham Young University. 894 p.
Welsh, S.L.; Atwood, N.D.; Goodrich, S.; Higgins, L.C., eds. 2015. A Utah Flora. Fifth Edition, revised. Provo, UT: Brigham Young University. 990 p.
West, N.E.; Reese, G.A. 1984. Average seasonal phytomass: A temporally independent index of herbaceous plant dominance. Vegetatio. 57(2-3): 137-141.
West, N.E.; Reese, G.A. 1991. Comparison of some methods for collecting and analyzing data on aboveground net production and diversity of herbaceous vegetation in a northern Utah subalpine context. Vegetatio. 96(2): 145-163.
Willms, W.D.; Quinton, D.A. 1995. Grazing effects on germinable seeds on the fescue prairie. Journal of Range Management. 48(5): 423-430.
Wilson, J.S.; Wilson, L.E.; Loftis, L.D.; Griswold, T. 2010. The montane bee fauna of north central Washington, USA, with floral associations. Western North American Naturalist. 70(2): 198-207.
Young, S.A.; Schrumpf, B.; Bouck, M.; Moore, M. 2020. How the Association of Official Seed Certifying Agencies (AOSCA) tracks wildland sourced seed and other plant propagating materials. Logan, UT: Utah State University, Utah Agricultural Experiment Station. 36 p.
How to Cite
Gucker, Corey L.; Shaw, Nancy L. 2025. Aspen fleabane (Erigeron speciosus). In: Gucker, C.L.; Shaw, N.L., eds. Western forbs: Biology, ecology, and use in restoration. Reno, NV: Great Basin Fire Science Exchange. Online: https://westernforbs.org