Authorship
Shaw, Nancy L.; Gucker, Corey L.
Publication Date
January 2018
Update
February 2024
Nomenclature
Sagebrush false dandelion (Nothocalais troximoides [A. Gray] Greene) (Greene 1886; Chambers 2006; GBIF 2024; ITIS 2024) is a member of family Asteraceae and tribe Cichorieae.
First Collections. Holotype: Microseris troximoides Gray (Gray 1884). Collected: Clear Water, Nez Perce County, Idaho by H.H. Spalding, Presbyterian missionary to the Nez Perce Tribe. Curated: Harvard University Herbaria (GBIF 2024).
Holotype: Nothocalais suksdorfii Greene (Greene 1886). Collected: western part of Klickitat County, Washington in 1882 by W.N. Suksdorf, botanist and plant collector. Curated: California Academy of Science (GBIF 2024).
Family
Asteraceae – Aster family
Genus
Nothocalais
Species
troximoides
NRCS Plant Code
NOTR2 (USDA NRCS 2024).
Synonyms
Microseris troximoides A. Gray, Nothocalais suksdorfii Greene, Scorzonella troximoides (A. Gray) Jepson (Greene 1886; Chambers 2006; ITIS 2024).
Common Names
Sagebush false dandelion, false agoseris, weevil false-dandelion, weevil prairie dandelion (Chambers 2006; GBIF 2024; USDA NRCS 2024).
Chromosome Number
The species is diploid with 2n = 18 (Stebbins et al. 1953; Chambers 2006).
Hybridization
Variability noted in Idaho, Montana, and Oregon, particularly on the lower Snake River Plain and in southeastern Oregon, may be the result of introgression with speckled false dandelion (N. nigrescens) and is typified by the presence of large involucres and phyllaries (Chambers 2006; Hitchcock and Cronquist 2018).
Distribution
Sagebrush false dandelion is distributed from south-central British Columbia to northern California, northern Nevada and Utah, largely east of the Cascade and Sierra Nevada mountains, and eastward to central Montana and northwestern Wyoming (CPNWH 2017; SEINet 2017; USGS 2017; USDA NRCS 2024).
Habitat And Plant Associations
Sagebrush false dandelion grows in dry to vernally moist sites from lowlands and foothills to mountainous areas (Cronquist 1955, 1994; Chambers 1993, 2006). The species is a component of intermountain grasslands, mountain grasslands, sagebrush (Artemisia spp.) steppe, sagebrush scrub, open pinyon-juniper (Pinus–Juniperus spp.) woodlands, northern juniper woodlands, northern oak (Quercus spp.) woodlands, and ponderosa pine (P. ponderosa) forests (Munz and Keck 1973; Pokorney et al. 2004; Chambers 2006; Kerns et al. 2006; Schuller and Halvorson 2009).
Elevation
Sagebrush false dandelion generally occurs at elevations from 160 to 6,600 ft (50-2,000 m) (Chambers 2006). A few collections have been made at elevations as great as 7,500 ft (2,300 m) in Idaho and 8,763 ft (2,671 m) in Nevada (SEINet 2017).
Soils
Sagebrush false dandelion grows on clayey to sandy loam soils of varied depths that are often stony (O’Farrell 1975; Pitt and Wikeem 1990; Sapsis 1990; Chambers 2006; Rhodes et al. 2010; Hitchcock and Cronquist 2018). It occurs in stiff sagebrush (A. rigida) communities of southeast Oregon on rocky scablands with very shallow to shallow stony soils derived from basalt or rhyolite. Soils in these communities may be saturated in winter and spring and subject to frost heaving (Ashley 2004).
Description
Sagebrush false dandelion is a somewhat variable acaulescent perennial herb with milky juice (Fig. 1) that resembles the common dandelion (Taraxacum officinale). The taproot is stout, fleshy, and covered with a blackish periderm. Lateral rootlets are borne on knoblike projections (Chambers 2006; GBIF 2024). One or more slender peduncles (flowering stems) 2 to 18 in (5-45 cm) tall arise from a thick caudex (Chambers 1993, 2006; Hitchcock and Cronquist 2018; GBIF 2024). Peduncles are erect, generally leafless, and glabrous. They are puberulent below the flower head, and may have 1 to 3 leafy bracts proximally (Chambers 2006; GBIF 2024). Leaves are basal, crowded, linear, lanceolate or occasionally divided, and usually 2 to 12 in (5-30 cm) long, less than 0.4 in (1 cm) wide, and acuminate (Cronquist 1955; Munz and Keck 1973; Taylor 1992; GBIF 2024). Leaf surfaces are glabrous or villous, particularly along the midveins. The margins are often wavy and white ciliolate (Cronquist 1955; Mansfield 2000; Chambers 2006; Welsh et al. 2015; GBIF 2024).


Figure 1. Sagebrush false dandelion plant in flower. Photos: M. Lavin, Montana State University.
Inflorescences (Fig. 2) are erect or nodding heads of 13 to 90 ligulate ray florets borne singly on each peduncle (Chambers 1993; Mansfield 2000; Welsh 2015). Flower heads are each subtended by an involucre 0.6 to 1 in (15-39 mm) wide consisting of 8 to 25 subequal, linear to lanceolate bracts (phyllaries) with the outer series shorter than or equal to the inner (Chambers 1993, 2006). The bracts are acuminate at the tip, green, sometimes with purple dots, and glabrous to white-hairy, especially on the margins and midrib, which is often purple (Cronquist 1955; Chambers 1993, 2006; Welsh 2015; Hitchcock and Cronquist 2018). The florets exceed the bracts (GBIF 2024). Ligules are 10 to 25 mm long, yellow, sometimes with an orange midrib that may become pinkish with age. Florets may close early in the day during hot weather (Taylor 1992; Chambers 1993, 2006).

 Figure 2. Sagebrush false dandelion flower heads with involucral bracts and ligulate ray florets. Photos: M. Lavin, Montana State University.
Figure 2. Sagebrush false dandelion flower heads with involucral bracts and ligulate ray florets. Photos: M. Lavin, Montana State University.
The fruit (a cypsela or false achene, often referred to as an achene) is cylindric or fusiform, narrowed and puberulent apically, 0.3 to 0.5 in (7-13 mm) long, beakless, and gray to light brown (Cronquist 1955; Chambers 1993, 2006). The pappus consists of 10 to 30 lustrous, white subulate or aristate scales (10-17 mm long) that taper to a scabrous bristle (Chambers 1993, 2006; Welsh 2015; GBIF 2024). Mature fruiting heads are white and spherical, resembling those of dandelions. Individual fruits resemble parachutes and are wind dispersed (Taylor 1992).
Reproduction
Sagebrush false dandelion reproduces entirely from seed.
Phenology
Flowering occurs from March to June depending on location (Chambers 2006). Pitt and Wikeem (1990) examined phenology of vegetation in a transitional zone between big sagebrush and ponderosa pine in south-central British Columbia at an elevation of 1,800 to 2,500 ft (550-750 m). They placed sagebrush false dandelion in a group of shallowly rooted, spring-ephemeral forbs that initiate development in very early spring, flower and complete growth rapidly to avoid summer drought, and rarely resprout in fall (Pitt and Wikeem 1990). In 1978 sagebrush false dandelion initiated growth and floral production in April, reached full flower in May, set seed and shattered in June, and plants were cured in July. In 1979, possibly due to lower April precipitation and soil moisture, seed set and shatter occurred in May, and plants cured in June (Pitt and Wikeem 1990).
Pollination
Pollination biology of sagebrush false dandelion has not been examined. Prairie false dandelion (N. cuspidata) is a hill and sand prairie species of the midwestern United States and the prairie provinces of Canada (Dieringer and Cabrera 2017). In Missouri it is primarily self-incompatible; most florets cross-pollinated over a 2- to 3-week period (Dieringer and Cabrera 2017). Bees considered likely pollinators were members of the Halictidae (4 species), Megachilidae (1 species), and Apidae (1 species).
Diseases
Podosphaera macularis (formerly Sphaerotheca macularis), powdery mildew of hops (Humulus lupulus), was identified on sagebrush false dandelion in Washington (Shaw 1973). This ascomycete infects species in many plant families, but its pathogenicity to sagebrush false dandelion has not been determined.
Ecology
Sagebrush false dandelion is a disturbance-tolerant forb that generally occurs as scattered plants within its communities. It can occur in early seral sites invaded by cheatgrass (Bromus tectorum) and on burned sites (Powell 1994; Wrobleski 1999). Following fire the species recovers by basal sprouting of surviving plants and establishment of windborne seed from non-burned areas (Powell 1994; Wrobleski 1999). Sagebrush false dandelion also persists in late-seral communities including Intermountain bunchgrass, sagebrush steppe, sagebrush scrub, ponderosa pine forests, pinyon-juniper woodland, and juniper woodland (Daubenmire 1992; Schuller and Halvorson 2009).
Disturbance Ecology
Fire response. Fall burns may not negatively impact sagebrush false dandelion (Cooper et al. 2011; Link and Hill n.d.), but decreased frequency following spring burns has been reported (Sapsis 1990).
Fall prescribed burns were conducted to determine whether they would increase the forb component in a Wyoming big sagebrush (Artemisia tridentata subsp. wyomingensis) community in the Hart Mountain Antelope Refuge in south-central Oregon. Six years following treatment there was no increase in yield of Cichorieae tribe forbs (sagebrush false dandelion, taper-tip hawksbeard [Crepis acuminata] and pale agoseris [Agoseris glauca]; P = 0.278) (Rhodes 2010). Another study conducted in this area found no significant differences in frequency, abundance, or density of sagebrush false dandelion between fall burned and non-burned plots in the first post-fire year (P > 0.10) (Wrobleski 1999).
Over a 3-year period following fall prescribed fires, frequency of sagebrush false dandelion increased from 0 to 80% on unburned control sites and from 20 to 100% on burned sites (Link and Hill n.d.). Sites were located in a cheatgrass-dominated area of a former big sagebrush/bluebunch wheatgrass (A. tridentata/Pseudoroegneria spicata) association (Daubenmire 1970) on the Columbia National Wildlife Refuge in central Washington.
Cooper et al. (2011) examined post-fire recovery of Wyoming big sagebrush steppe on 24 sites in central and southeastern Montana 4 to 67 years following prescribed or natural fires. Members of the Asteraceae, tribe Chichorieae, were expected to increase following fire. Native Chichorieae, sagebrush false dandelion and pale agoseris, were very rare with a fewer than 40 plants occurring in plots totaling 15,500 ft² (1,440 m²). When combined with non-native Chichorieae, yellow salsify (Tragopogon dubius) and common dandelion, overall density was similar for burned and unburned areas (P > 0.05).
Sagebrush false dandelion may be sensitive to spring burning. Its frequency decreased from 20 to 4% in the second season following prescribed spring fires, but was unchanged following fall prescribed burns (P > 0.10) (Sapsis 1990). Burns were conducted in a basin big sagebrush (A. tridentata subsp. tridentata)/ perennial bunchgrass community in the John Day Fossil Beds National Monument in eastern Oregon.
In low sagebrush (Artemisia arbuscula var. arbuscula) communities in southeastern Oregon, density of several tall forb species including sagebrush false dandelion (P<0.001) increased by 1.4 to 2.9 times compared to non-burned sites eight years after fall prescribed burns (Bates and Davies 2022). Although differences were not significant, yield of tall forbs in the burned sites averaged 137 lbs/ac (154 kg/ha) greater than in the non-burned sites.
Wildlife And Livestock Use
Sagebrush false dandelion use or association with key habitat has been reported for pocket gophers (Thomomys talpoides) (O’Farrell 1975), Great Basin pocket mice (Perognathus parvus) (Richardson et al. 2013), greater sage-grouse (Centrocercus urophasianus) (Wrobleski 1999; Wik 2002; Rhodes et al. 2010), and blue grouse (Dendragraphus obscurus) (Galvin 1979).
Small mammals. O’Farrell (1975) examined pocket gopher abundance in seven vegetation associations on the Hanford Reserve Arid Lands Ecology Site in central Washington. Greatest abundance occurred in abandoned wheat fields invaded by cheatgrass and tall tumblemustard (Sisymbrium altissimum) within the sagebrush/wheatgrass association. It was hypothesized that pocket gopher use of sagebrush false dandelion and other tap-rooted species present in these fields might have concentrated their populations on these sites relative to undisturbed sagebrush/wheatgrass stands.
Birds. Many studies indicate that seeds of forbs in the chicory tribe of the sunflower family are favored in summer diets of greater sage-grouse and are important for successful brood rearing (Barnett and Crawford 1994, Drut et al. 1994; Dumroese et al. 2015; Stiver et al. 2015). Sagebrush false dandelion was one of the forbs in intact Wyoming big sagebrush steppe of the northern Great Basin of southcentral Oregon known to be used by greater sage-grouse (Rhodes et al. 2010). At the Hart Mountain Antelope Refuge in this area it was identified as 1 of 19 species prevalent in greater sage-grouse diets (Wrobleski 1999). In south-central Owyhee County, Idaho, sagebrush false dandelion was found at greater sage-grouse brood-rearing sites (Wik 2002).
Galvin (1979) reported that the flowers of sagebrush false dandelion were eaten by blue grouse in wheatgrass/bluegrass communities of the Palouse prairie in eastern Washington.
Ethnobotany
Tissues of sagebrush false dandelion contain a bitter, milky juice that the Plains Indians may have chewed as gum (Taylor 1992).
Revegetation Use
Sagebrush false dandelion is of interest as a revegetation species because it is fire tolerant, used by greater sage-grouse, and spreads from windblown seed (Sapsis 1990; Powell 1994; Wrobleski 1999; Rhodes et al. 2010). To date it has received limited use in wildland plantings. Evaluating its potential as a revegetation species necessitates examination of its seed production in agricultural fields and identification of techniques for establishing it on wildland sites.
The seeds of sagebrush false dandelion are not removed from the fruit (cypsela or false achene) during the cleaning process. Consequently, the term “seed” as used by collectors, growers, and users refers to the fruits and that convention is applied in the following sections.
Developing A Seed Supply
For restoration to be successful, the right seed needs to be planted in the right place at the right time. Coordinated planning and cooperation is required of all partners to select appropriate species and seed sources; these are determined by current or projected revegetation requirements and goals (PCA 2015).
Developing a seed supply begins with seed collection from native stands. Collection sites are determined by current or projected revegetation requirements and goals as well as through consultation with land managers and stakeholders. Collected seed is distributed for restoration or first increased in seed fields to provide a larger supply of seed (PCA 2015). Production of nursery stock requires less seed than large-scale seeding operations. Regardless of the size and complexity of any revegetation effort, seed certification is essential for tracking seed origin from collection through use (UCIA 2015).
Seed Sourcing
Because empirical seed zones are not currently available for sagebrush false dandelion, generalized provisional seed zones developed by Bower et al. (2014), may be used to select and deploy seed sources. These provisional seed zones identify areas of climatic similarity with comparable winter minimum temperature and aridity (annual heat:moisture index). In Figure 3, Omernik Level III Ecoregions (Omernik 1987) overlay the provisional seed zones to identify climatically similar but ecologically different areas. For site-specific disturbance regimes and restoration objectives, seed collection locations within a seed zone and ecoregion may be further limited by elevation, soil type, or other factors.
The Western Wildland Environmental Threat Assessment Center’s Threat and Resource Mapping (TRM) Seed Zone application (USFS WWETAC 2017) provides links to interactive mapping features useful for seed collection and deployment planning. The Climate Smart Restoration Tool (Richardson et al. 2020) can also guide revegetation planning, seed collection, and seed deployment, particularly when addressing climate change considerations.
Occurrence Map
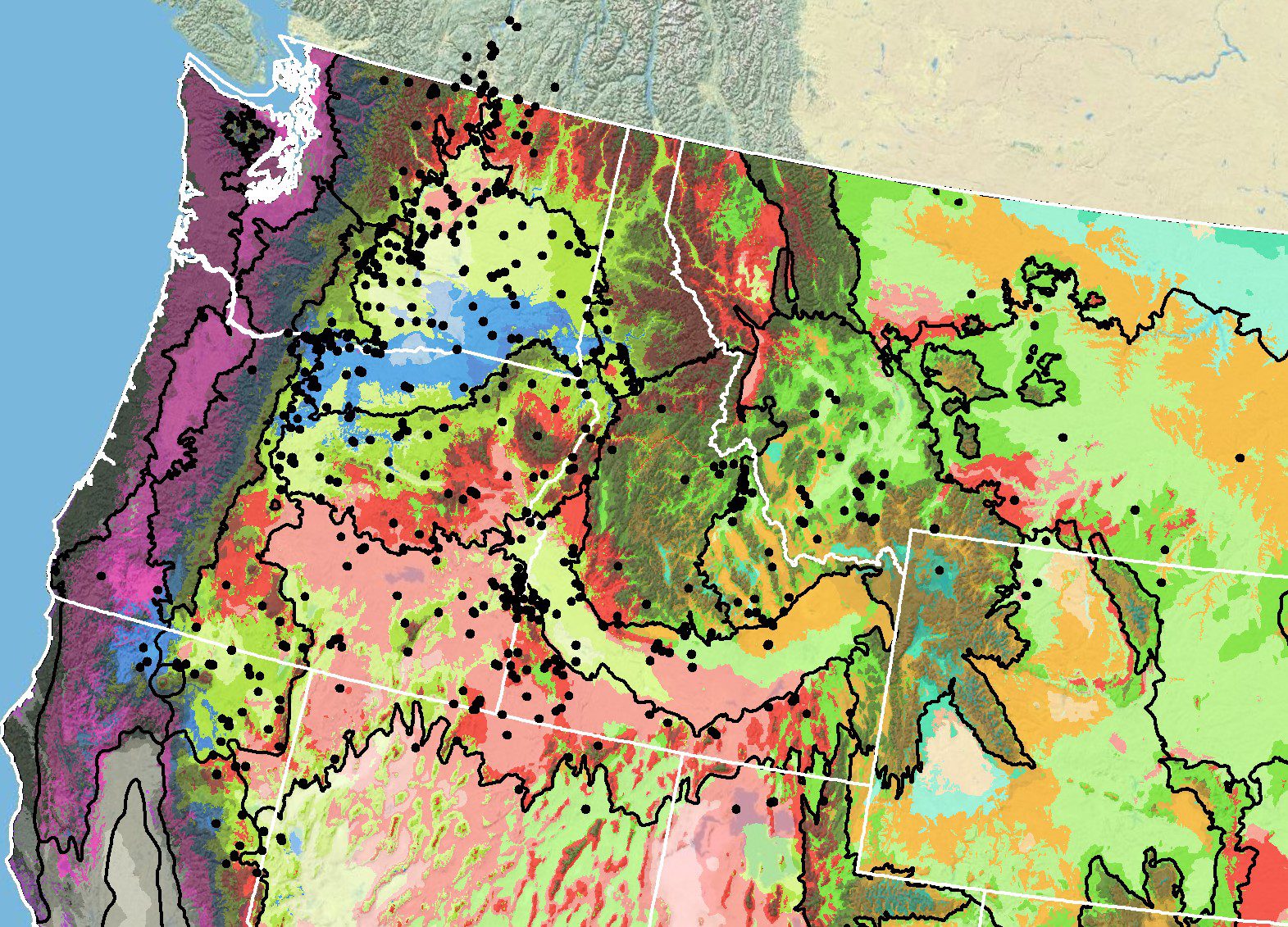
Figure 3. Distribution of sagebrush false dandelion (black circles) based on geo-referenced herbarium specimens and observational data from 1876-2016 (dates not available for all points) (CPNWH 2017; SEINet 2017; USGS 2017). Generalized provisional seed zones (colored regions; Bower et al. 2014) are overlain by Omernik Level III Ecoregions (black outlines) (Omernik 1987; USDI EPA 2017). Interactive maps, legends, and a mobile app are available (USFS WWETAC 2024). Map prepared by M. Fisk, USGS.
Releases
There are currently (early 2024) no released sagebrush false dandelion germplasms.
Wildland Seed Collection
Sagebrush false dandelion plants vary in overall size and production of seedheads, depending on location and variation in annual precipitation (B. Youtie and S. Jackman, Eastern Oregon Stewardship Services [EOSS]), personal communications, August 2017). Plants may be small and produce a single seed head on dry sites or in dry years, or they may be larger with multiple peduncles and seed heads on more mesic areas or in wet years (Fig. 4). Seed heads ripen over a fairly short time period, usually about 2 to 4 weeks, but this can be shortened by wind and rain (B. Youtie and S. Jackman, EOSS, personal communications, August 2017).


Figure 4. Sagebrush false dandelion plants with mature and dispersing seedheads illustrating uneven ripening. Photos: M. Lavin, Montana State University.
Wildland Seed Certification
Verification of species and tracking of geographic source is necessary whether wildland seed is collected for immediate project use or as stock seed for cultivated increase. This official Source Identification process can be accomplished by following procedures established by the Association of Official Seed Certifying Agencies (AOSCA) Pre-Variety Germplasm Program (UCIA 2015; Young et al. 2020). Wildland seed collectors should become acquainted with state certification agency procedures, regulations, and deadlines in the states where they collect.
If wildland-collected seed is to be sold for direct use in ecological restoration projects, collectors must apply for Source-Identified certification prior to making collections. Pre-collection applications, site inspections, and species and seed amount verification are handled by the AOSCA member state agency where seed collections will be made (see listings at AOSCA.org).
If wildland seed collected by a grower or private collector is to be used as stock seed for planting cultivated seed fields or for nursery propagation (See Agricultural Seed Field Certification section), detailed information regarding collection site and collecting procedures must be provided when applying for certification. Photos and herbarium specimens may be required. Germplasm accessions acquired within established protocols of recognized public agencies, however, are normally eligible to enter the certification process as stock seed without routine certification agency site inspections. For contract grow-outs, however, this collection site information must be provided to the grower to enable certification.
Collection Timing
Camp and Sanderson (2007) collected seed in Washington at elevations from 1,830 to 2,260 ft (560-690 m) between late May and early June. Collection dates for 21 Seeds of Success seed lots ranged from April 30 in 2013 to June 24 in 2011 with the collections from the earliest and latest dates both from Malheur County, Oregon (elevations not provided) (USDI BLM SOS 2017, 2024). Collection date was May 19, 2010 for the lowest recorded elevation site (2,000 ft [600 m]) in Douglas County, Washington, and June 15, 2020 for the highest recorded elevation (7,939 ft [2,420 m]) in Johnson County, Wyoming (USDI BLM SOS 2017, 2024).
The window for seed collection lasts about 7 to 10 days, even though seeds are present over a longer period (Camp and Sanderson 2007). Collection near the mid-point of the seed maturation period permits harvest of more early, mid-, and late-maturing seed if it is not possible to collect on multiple dates. Camp and Sanderson (2007) reported that most seed heads ripened uniformly in eastern Washington, while B. Youtie and S. Jackman (EOSS, personal communication, August 2017) found flowering to be somewhat uneven among plants within populations and among seed heads on individual plants in eastern Oregon (Fig 4).
Collection Methods
Seed collection of sagebrush false dandelion is time consuming and large harvests are rare. Plants are generally scattered, seeds must be harvested manually, and per plant production is low. Seed quality can be estimated prior to collection using cut tests or X-ray tests. Seeds are collected manually by plucking them from individual seed heads that are white, expanded, and spherical (Camp and Sanderson 2007). Seeds are peacock green when immature and brown at maturity (Chambers 1993, 2006; Camp and Sanderson 2007). Only seeds that dislodge easily should be collected to avoid harvesting insect- or disease-damaged seeds. Seed heads with mature-appearing seeds clumped together may be infested with weevils (Camp and Sanderson 2007). Clipping is an alternative if seed heads are held over a container and there is no wind.
Species of the false dandelion (Nothocalais spp.) and agoseris (Agoseris spp.) genera can be difficult to distinguish when plants have dried as their immature fruits are similar in appearance. The ranges of the two species overlap, and fruits of both species mature at about the same time. False dandelion species have a white to silvery pappus, while that of agoseris species is creamy and soft. Fruits of false dandelion lack the beak present in agoseris fruits (Fig. 5; Cronquist 1994; B. Youtie, EOSS, personal communication, August, 2017).
Several collection guidelines and methods should be followed to maximize the genetic diversity of wildland collections: 1) collect seed from a minimum of 50 randomly selected plants; 2) collect from widely separated individuals throughout a population without favoring the most robust or avoiding small stature plants; and 3) collect from all microsites including habitat edges (Basey et al. 2015). General collecting recommendations and guidelines are provided in online manuals (e.g., ENSCONET 2009; USDI BLM SOS 2023).
It is critical that wildland seed collection does not impact the sustainability of native plant populations. Collectors should take no more than 20% of the viable seed available at the time of harvest (USDI BLM SOS 2023). Additionally, care must be taken to avoid the inadvertent collection of weedy species, particularly those that produce seeds similar in shape and size to those of false dandelion.
Post-Collection Management
Molding can occur if leaves and stems, which contain milky juice, are collected along with the seeds, particularly if collected during high humidity periods. Vegetative material should be removed promptly by hand or with sieves. Harvested seed should be spread on racks in a protected area and thoroughly air dried in the field or following transport to the cleaning facility. Collected material can be transported in clean, breathable bags or boxes and should be protected from overheating during transport. Insect infestations should be controlled by freezing collections for 48 hours or use of appropriate chemicals. Temporary storage prior to delivery to the seed extractory should be in a cool (<50 ºF [10 ºC]), dry (<35% relative humidity) location.


Figure 5. Mature fruits (cypselas, false achenes) of sagebrush false dandelion with pappus bristles. Note the lack of beaks on the fruits. Photos: M. Lavin, Montana State University (top), BLM ID931 SOS (bottom, scale: mm).
Seed Cleaning
Seed quality of bulk seed lots can be estimated using X-ray or cut tests. An office clipper, screens, or sieves can be used to remove large inert material (Fig. 6). Pappus removal is accomplished by hand working the material on a rubbing board or running it through a dewinger. Pappus material and other debris are removed using a seed clipper or a seed blower, depending on seed lot size.
Seed Storage
Seeds of sagebrush false dandelion are orthodox. Seeds frozen for 2 months at the Royal Botanic Garden-Kew at -4 ºF (-20 ºC) and 15% relative humidity retained 100% viability (SER, INSR, RBGK 2023).
Seed Testing
There is no AOSA rule for testing germination or protocol for examining viability of sagebrush false dandelion (AOSA 2016). Purity may be evaluated using standard AOSA procedures.


Figure 6. Sagebrush false dandelion seed after initial cleaning (above), and seed following final cleaning (below) at the USFS Bend Seed Extractory. Photos: USFS.
Germination Biology
Germination of sagebrush false dandelion seed was tested at the Royal Botanic Gardens – Kew (WP) by germinating untreated seed on 1% agar at temperatures ranging from 50 to 77 oF (10 to 25 oC) with exposure to an 8 hr light/16 hr dark regime. Germination exceeded 90% at all temperatures tested. Time to final germination ranged from 56 days at 68 °F (20oC) to 231 days at 59 °F (15oC) (SER, INSR, RBGK 2023).
Wildland Seed Yield And Quality
The cleanout ratio (clean seed weight:rough weight of seed collection) for seed lots of sagebrush false dandelion cleaned at the USFS Bend Seed Extractory and seed quality data for these lots are provided in Table 1) . Purity and viability of cleaned seed lots are often above 90% (USFS BSE 2017, 2023). The number of seeds per pound reported by other sources falls within the range reported in Table 1. The Seed Information Database reported an average of 158,902 seeds/lb (350,319 seeds/kg) for two lots (SER, INSR, RGBK 2023), while Camp and Sanderson (2007) reported a BLM count of 204,300 seeds/lb (450,400 seeds/kg).
Table 1. Seed yield and quality of sagebrush false dandelion seed lots collected in the Intermountain region, cleaned and evaluated by the Bend Seed Extractory. Viability was tested by the Oregon State Seed Laboratory or the USFS National Seed Laboratory (USFS BSE 2017, 2023).
|
Characteristic |
Mean |
Range |
Samples (#) |
|
Bulk weight (lbs) |
0.56 |
0.07-5.13 |
51 |
|
Clean weight (lbs) |
0.125 |
0.01-2.82 |
51 |
|
Clean-out ratio |
0.36 |
0.13-0.64 |
51 |
|
Purity (%) |
98 |
92-99 |
51 |
|
Fill (%)¹ |
90 |
70-99 |
51 |
|
Viability (%)² |
93 |
88-97 |
16 |
|
Seeds/lb |
166,524 |
77,538-396,572 |
51 |
|
Pure live seeds/lb |
162,056 |
71,389-369,050 |
16 |
¹100 seed X-ray test
²Tetrazolium chloride test
Marketing Standards
Acceptable seed purity, viability, and germination specifications vary with revegetation plans. High purity and viability are required for precision seeding equipment used in nurseries and agricultural seed fields. Lower purity may be acceptable for some wildland seeding techniques.
Agricultural Seed Production
Sagebrush false dandelion has been grown for seed production under contract, but a production protocol is not available. The USDI Bureau of Land Management ID/IQ (indefinite delivery/indefinite quantity) Contract (USDI BLM NOC 2017) lists a seeding rate of 5 lbs pure live seed (PLS)/ac (5.6 PLS kg/ha) and a potential seed yield of 150 lbs/ac (170 kg/ha).
Agricultural Seed Certification
In order to minimize genetic changes in specific accessions of native species when increased in cultivated fields, it is essential to track the geographic source and prevent inadvertent hybridization or selection pressure. This is accomplished by following third party seed certification protocols for Pre-Variety Germplasm (PVG) as established by the Association of Official Seed Certification Agencies (AOSCA). AOSCA members in the U.S., Canada, and other countries administer PVG requirements and standards that track the source and generation of planting stock. Field and cleaning facility inspections then monitor stand establishment, proper isolation distances, control of prohibited weeds, seed harvesting, cleaning, sampling, testing, and labeling for commercial sales (UCIA 2015; Young et al. 2020).
Seed growers apply for certification of their production fields prior to planting and plant only certified stock seed of an allowed generation (usually less than four). The systematic and sequential tracking through the certification process requires preplanning, knowing state regulations and deadlines, and is most smoothly navigated by working closely with state certification agency personnel. See the Wildland Seed Certification section for more information on stock seed sourcing.
Nursery Practice
Methow Natives, Winthrop, Washington, propagated sagebrush false dandelion in flats using a 1:1:1 soil mix of peat moss, perlite and vermiculite (BLM Forb Propagation Notes 2003–2006 reviewed by Camp and Sanderson 2007). The soil mix was amended with dolomite lime, NPK 8-6-12 fertilizer, superphosphate, and micronutrient fertilizer. Germination occurred if untreated seeds were planted in spring without stratification, but not when seeds were stratified outdoors for 4 months in winter before being moved into a greenhouse. Seedlings were planted in 10 in³ (164 cm³) cone-tainers and held over the summer for fall planting.
Wildland Seeding And Planting
No guidelines for large-scale seeding of sagebrush false dandelion are available. In a small-plot seeding trial, 10 to 20 seeds were fall planted per plot yielding at least one plant per plot the next spring (Camp and Sanderson 2007).
In a second small-plot planting trial, seedlings were planted in fall with or without irrigation. Survival was 42% for irrigated and 45% for non-irrigated seedlings (monitoring date and planting location not provided) (Camp and Sanderson 2007). Container seedlings may be planted in fall or spring (USDI BLM NOC 2017).
Weed control. Sagebrush false dandelion was among 61 dicot species found to be resistant to Pseudomonas fluorescens strains ACK55, NKK78, and SMK69 that inhibit the annual weedy grasses cheatgrass, jointed goatgrass (Aegilops cylindrica), and medusahead (Taeniatherum caput-medusae) (Kennedy 2018). These strains were also selected for traits that minimize negative effects on microbial populations and other organisms. Further testing will aim to identify strains that are more effective in specific conditions.
Monitoring. Because little information is available to guide planting and seeding projects that include sagebrush false dandelion, long-term monitoring is essential to evaluate seeding or planting success and inform future projects.
Acknowledgements
Funding for Western Forbs: Biology, Ecology, and Use in Restoration was provided by the USDI BLM, Great Basin Ecoregional Program through the Great Basin Fire Science Exchange. We thank Berta Youtie and Siri Jackman for reviewing the chapter and providing seed collection recommendations.
This research was supported in part by the USDA Forest Service, Rocky Mountain Research Station. The findings and conclusions in this publication are those of the authors and should not be construed to represent any official USDA or U.S. Government determination or policy.
Literature Cited
Ashley, P. 2004. Malheur River Wildlife Mitigation Project. 2003 Annual Report, Project 200002700. Doe/bp-00004050-1. Portland, OR: Bonneville Power Administration. 208 p. http://bptdnr.com/wp-content/uploads/2016/08/BPT_Wildlife_AnnualReport_Malheur_2003.pdf [Accessed 2017 December 28].
Association of Official Seed Analysts [AOSA]. 2016. AOSA rules for testing seeds. Vol. 1. Principles and procedures. Washington, DC: Association of Official Seed Analysts.
Barnett, J.K.; Crawford, J.A. 1994. Pre-laying nutrition of sage grouse hens in Oregon. Journal of Range Management. 47(2): 114-118.
Basey, A.C.; Fant, J.B.; Kramer, A.T. 2015. Producing native plant materials for restoration: 10 rules to collect and maintain genetic diversity. Native Plants Journal. 16(1): 37-53.
Bates, J.D.; Davies, K.W. 2022. Early succession following prescribed fire in low sagebrush (Artemisia arbuscula var. arbuscula) steppe. 82(1): 50-66.
Bower, A.D.; St. Clair, J.B.; Erickson, V. 2014. Generalized provisional seed zones for native plants. Ecological Applications. 24(5): 913-919.
Camp, P.; Sanderson, J. 2007. Seed collection, propagation and reintroduction of native wildflowers in the Columbia Basin. Wenatchee, WA: U.S. Department of the Interior, Bureau of Land Management, Wenatchee Field Office. 32 p.
Chambers, K.L. 1993. Nothocalais False-agoseris. In: Hickman, J.C, ed. The Jepson manual: Higher plants of California. Berkeley, CA: University of California Press. 320 p.
Chambers, K.L. 2006. 67. Nothocalais. In: Flora of North America Editorial Committee, eds. Flora of North America North of Mexico. Volume 19 Magnoliophyta: Asteridae, part 6: Asteraceae, part 1, Asterales, part 1 (Aster order). New York, NY: Oxford University Press: 335-337.
Consortium of Pacific Northwest Herbaria [CPNWH]. 2017. Nothocalais troximoides. Seattle, WA: University of Washington Herbarium, Burke Museum of Natural History and Culture. pnwherbaria.org/ [Accessed 2017 September 18].
Cooper, S.V.; Lesica, P.; Kudray, G.M. 2011. Post-fire recovery of Wyoming big sagebrush steppe in central and southeast Montana. Natural Resources and Environmental Issues. 16(12): 1-9.
Global Biodiversity Information Facility [GBIF]. 2024. GBIF Secretariat. Available: https://www.gbif.org/
Hitchcock, C.L.; Cronquist, A. 2018. Flora of the Pacific Northwest: An illustrated manual. Second Ed. Giblin, D.E.; Legler, B.S.; Zika, P.F.; Olmstead, R.G., eds. Seattle, WA: University of Washington Press. 882 p.
ITIS Database. 2024. Integrated Taxonomic Information System. Available: http://www.itis.gov/index.html
Mansfield, D.H. 2000. Flora of Steens Mountain. Corvallis, OR: Oregon State University Press. 410 p.
Munz, P.A.; Keck, D.D. 1973. A California flora and supplement. Berkeley, CA: University of California Press. 1905 p.
O’Farrell, T.P. 1975. Small mammals, their parasites and pathologic lesions on the Arid Lands Ecology Reserve, Benton County, Washington. The American Midland Naturalist. 93(2): 377-387.
Omernik, J.M. 1987. Ecoregions of the conterminous United States. Map (scale 1:7,500,000). Annals of the Association of American Geographers. 77(1): 118-125.
Pitt, M.D.; Wikeem, B.M. 1990. Phenological patterns and adaptations in an Artemisia/Agropyron plant community. Journal of Range Management. 43(4): 350-358.
Pokorney, M.L.; Sheley, R.L.; Svejcar, T.J.; Engel, R.E. 2004. Plant species diversity in a grassland plant community: Evidence for forbs as a critical management consideration. Western North American Naturalist. 64(2): 219-230.
Powell, D.C. 1994. Effects of the 1980s western spruce budworm outbreak on the Malheur National Forest in Northeastern Oregon. Technical Publication R6-FI&D-TP-12-94. Portland, OR: U.S. Department of Agriculture, Forest Service, Natural Resources Staff, Forest Insects and Diseases Group. 176 p.
Rhodes, E.C.; Bates, J.D.; Sharp, R.N.; Davies, K.W. 2010. Fire effects on cover and dietary resources of sage-grouse habitat. The Journal of Wildlife Management. 74(4): 755-764.
Richardson, B.; Kilkenny, F.; St. Clair, B.; Stevenson-Molnar, N. 2020. Climate Smart Restoration Tool. https://climaterestorationtool.org/csrt/
Sapsis, D.B. 1990. Ecological effects of spring and fall prescribed burning on basin big sagebrush/Idaho fescue-bluebunch wheatgrass communities. Corvallis, OR: Oregon State University. Thesis. 106 p.
Schuller, R.; Halvorson, R. 2009. Benjamin Research Natural Area: Guidebook supplement 36. Gen. Tech. Rep. PNW-GTR-786. Portland, OR: U.S. Department of Agriculture, Forest Service, Pacific Northwest Research Station. 20 p.
SEINet – Regional Networks of North American Herbaria Steering Committee [SEINet]. 2017. SEINet Regional Networks of North American Herbaria. https://symbiota.org/seinet
Shaw, C.G. 1973. Host fungus index for the Pacific Northwest – I. Hosts. Washington State University Agricultural Experiment Station Bulletin. 765: 1-121.
Society for Ecological Restoration; International Network for Seed Based Restoration; Royal Botanic Gardens [SER, INSR, RBGK]. 2023. Seed Information Database (SID). Available from: https://ser-sid.org/
Stebbins, G.L., Jr.; Jenkins, J.A.; Walters, M.S. 1953. Chromosomes and phylogeny in the Compositae, Tribe Cichorieae. In: Foster, A.S.; Constance, L.; Papenfuss, G.F. Berkeley, CA: University of California Press. University of California Publications in Botany. 26(6): 401-430.
Stiver, S.J.; Rinkes, E.T.; Naugle, D.E.; Makela, P.D.; Nance, D.A.; Karl, J.W., eds. 2015. Sage-grouse habitat assessment framework: A multi-scale assessment tool. Technical Reference 6710-1. Denver, CO: U.S. Department of the Interior, Bureau of Land Management and Western Association of Fish and Wildlife Agencies. 114 p.
Taylor, R.J. 1992. Sagebrush country: A wildflower sanctuary. Missoula, MT: Mountain Press Publishing Company. 211 p.
USDA Forest Service, Bend Seed Extractory [USDA FS BSE]. 2017. Nursery Management Information System Version 4.1.11. Local Source Report 34-Source Received. Bend, OR: U.S. Department of Agriculture, Forest Service, Bend Seed Extractory.
USDA Forest Service, Bend Seed Extractory [USDA FS BSE]. 2023. Nursery Management Information System Version 4.1.11. Local Source Report 34-Source Received. Bend, OR: U.S. Department of Agriculture, Forest Service Bend Seed Extractory.
USDA Forest Service, Western Wildland Environmental Threat Assessment Center [USFS WWETAC]. 2017. TRM Seed Zone Applications. Prineville, OR: U.S. Department of Agriculture, Forest Service, Western Wildland Environmental Threat Assessment Center. https://research.fs.usda.gov/pnw/products/dataandtools/seed-zone-webmap
USDA Natural Resources Conservation Service [USDA NRCS]. 2024. The PLANTS Database. Greensboro, NC: U.S. Department of Agriculture, Natural Resources Conservation Service, National Plant Data Team. https://plants.sc.egov.usda.gov/
USDI Bureau of Land Management, National Operations Center [USDI BLM NOC]. 2017. FY2017 Native seed increase and supply. Appendix I and II. Denver, CO: U.S. Department of the Interior, Bureau of Land Management, National Operations Center.
USDI Bureau of Land Management, Seeds of Success [USDI BLM SOS]. 2017. Seeds of Success collection data. Washington, DC: U.S. Department of the Interior, Bureau of Land Management, Plant Conservation and Restoration Program.
USDI Bureau of Land Management, Seeds of Success [USDI BLM SOS]. 2023. Bureau of Land Management technical protocol for the collection, study, and conservation of seeds from native plant species for Seeds of Success. Washington, DC: U.S. Department of the Interior, Bureau of Land Management, Plant Conservation and Restoration Program. 45 p.
USDI Bureau of Land Management, Seeds of Success [USDI BLM SOS]. 2024. Seeds of Success collection data. Washington, DC: U.S. Department of the Interior, Bureau of Land Management, Plant Conservation and Restoration Program.
USDI Environmental Protection Agency [USDI EPA]. 2017. Ecoregions. Washington, DC: U.S. Department of the Interior, Environmental Protection Agency. https://www.epa.gov/eco-research/ecoregions
USDI Geological Survey [USDI USGS]. 2017. Biodiversity Information Serving Our Nation (BISON). U.S. Geological Survey. https://www.gbif.us/
Utah Crop Improvement Association [UCIA]. 2015. How to be a seed connoisseur. Logan, UT: UCIA, Utah Department of Agriculture and Food, Utah State University and Utah State Seed Laboratory. 16 p.
Welsh, S.L. 2015. Compositae Giseke Sunflower family. In: Welsh, S.L.; Atwood, N.D.; Goodrich, S.; Higgins, L.C., eds. and contributors. Fifth Ed. Provo, UT: Brigham Young University: 137-273.
Wik, P.A. 2002. Ecology of greater sage-grouse in south-central Owyhee County, Idaho. Moscow, ID: University of Idaho. Thesis. 141 p.
Wrobleski, D. 1999. Effects of prescribed fire on Wyoming big sagebrush communities: Implications for ecological restoration of sage grouse habitat. Corvallis, OR: Oregon State University. Thesis. 76 p.
Young, S.A.; Schrumpf, B.; Bouck, M.; Moore, M. 2020. How the Association of Official Seed Certifying Agencies (AOSCA) tracks wildland sourced seed and other plant propagating materials. Logan, UT: Utah State University, Utah Agricultural Experiment Station. 36 p.
How to Cite
Shaw, Nancy L.; Gucker, Corey L. 2024. Sagebrush false dandelion (Nothocalais troximoides). In: Gucker, C.L.; Shaw, N.L., eds. Western forbs: Biology, ecology, and use in restoration. Reno, NV: Great Basin Fire Science Exchange. Online: https://westernforbs.org