Authorship
Gucker, Corey; Shaw, Nancy
Publication Date
September 2018
Update
November 2025
Nomenclature
Sphaeralcea parvifolia A. Nelson is commonly referred to as small-leaf globemallow and is part of the the Malvaceae or Mallow family and the Eusphaeralcea subgenus (Kearney 1935).
Family
Malvaceae – Mallow family
Genus
Sphaeralcea
Species
parvifolia
NRCS Plant Code
SPPA2 (USDA NRCS 2025).
Subtaxa
The Flora of North America does not currently recognize any varieties or subspecies (La Duke 2016).
Synonyms
Sphaeralcea arizonica A. Heller ex Rydberg, S. marginata York ex Rydberg, S. ambigua subsp. rugosa Kearney, S. ambigua var. rugosa (Kearney) Kearney (Holmgren et al. 2005; La Duke 2016; ITIS 2025).
Common Names
Small-leaf globemallow, small-flower globemallow, Nelson globemallow (Vernon et al. 2005; LBJWC 2018; USDA NRCS 2025).
Chromosome Number
Chromosome numbers are: 2n = 10, 20 (La Duke 2016).
Hybridization
There is frequent hybridization and polyploidy within the Sphaeralcea genus, which results in indefinite and overlapping species boundaries and can make accurate identification difficult in some areas (Kearney 1935; La Duke 2016). Hybrids between small-leaf globemallow and Munro’s globemallow are common (Kearney 1935).
Distribution
Small-leaf globemallow occurs in the Central Basin and Range and Colorado Plateau ecoregions (Lambert 2005). It occurs naturally in Nevada, the southern two-thirds of Utah, western Colorado, northwestern New Mexico, northern and central Arizona, and adjacent California (Kearney 1935; Holmgren et al. 2005; La Duke 2016; USDA NRCS 2025). On the high plateaus of central and northern Arizona, small-leaf globemallow is the most abundant globemallow species (Kearney 1935). Although the distributions of small-leaf and Munro’s globemallow overlap in Nevada, Utah, and Colorado, it is common to find small-leaf globemallow in more southern and Munro’s globemallow in more northern locations (Sriladda et al. 2012).
Specimens of small-leaf globemallow collected in Oneida County, Idaho, in 2021 and Cache County, northern Utah, in 1993 represent persistent populations from restoration plantings (Daines et al. 2022) planted near these locations in 1987 by Pendery and Rumbaugh (1990).
Habitat And Plant Associations
Dry slopes, mesas, washes, roadsides, and disturbed habitats in warm desert shrub, salt desert shrub, sagebrush (Artemisia spp.), and mountain brush communities are common small-leaf globemallow habitats (Munz and Keck 1973, Holmgren et al. 2005, La Duke 2016; LBJWC 2018; Rink et al. 2020). Associated species in these habitats include greasewood (Sarcobatus spp.), saltbush (Artiplex spp.), blackbrush (Coleogyne ramosissima), and pinyon-juniper woodlands (Pinus–Juniperus spp.) (Fig. 1) (Munz and Keck 1973; Holmgren et al. 2005; Welsh et al. 2015).
When black sagebrush (Artemisia nova) vegetation was monitored for 4 yrs in western Nevada’s Buckskin Mountains, frequency of small-leaf globemallow was 3% in a dry spring with no moisture, but it was not sampled in a wet spring with above-average precipitation. Non-shrub vegetation was dominated by eastern bottlebrush grass (Elymus hystrix) in dry years and cheatgrass (Bromus tectorum) in wet years (Young and Palmquist 1992).

Figure 1. Small-leaf globemallow growing in Colorado. Photo: USDI Bureau of Land Management (BLM) CO150A, Seeds of Success (SOS).
Globemallows (small-leaf, desert [S. ambigua], gooseberryleaf [S. grossulariifolia], and Rusby’s [S. rusbyi]) were common in vegetation on the rim of Montezuma Well (an artesian fed lake) and in a natural limestone sink in central Arizona. These species occurred in a quarter of the plant communities dominated by broom snakeweed (Gutierrezia sarothrae) and mariola (Parthenium incanum) on southern exposures, with cover reaching a high of 0.7% (Clark and Burgess 1966). In a study of climate and vegetation patterns in vegetation representing a transition between Mojave and Great Basin deserts at the Nevada Test Site north of Las Vegas, small-leaf globemallow occurred with minor cover (0.1%) in vegetation dominated by Shockley’s desert-thorn (Lycium shockleyi). It did not occur in communities dominated by creosote bush (Larrea tridentata). Shrub and herbaceous cover was low in this area. Total cover of herbaceous perennials was 0.9% in Shockley’s desert-thornand 1.9% in creosote bush shrublands (Beatley 1975).
Elevation
Although it occurs at elevations of 2,000 to 9,000 ft (600-2,700 m) (Holmgren et al. 2005; La Duke 2016), small-leaf globemallow is rare below 4,000 ft (1,200 m) (Kearney 1935). In California, the elevation range for small-leaf globemallow is 5,000 to 7,000 ft (1,500-2,100 m) (Munz and Keck 1973), and in Utah it is 2,700 to 8,900 ft (820-2,700 m) (Welsh et al. 2015).
Soils
Small-leaf globemallow is often found in disturbed habitats with dry, gravelly, rocky, well-drained soils with sandy to clay texture (Kearney 1935; Jameson et al. 1962; Dorn and Dorn 2007; Cooney and Dreesen 2018; LBJWC 2023). The species occurs on sites having at least 6 in (15 cm) of soil (Jameson et al. 1962; Schmutz et al. 1967).
On the Hopi Indian Reservation in northeastern Arizona, small-leaf globemallow made up 1% of the species composition on coppice dunes but did not occur in dune interspaces. Soils at both locations were loamy fine sands, but the depth to sandstone was 12 to 36 in (30-91 cm) on the dunes and 3 to 5 in (8-13 cm) in the interspaces. Cutler’s jointfir (Ephedra cutleri) dominated the dunes and Indian ricegrass (Achnatherum hymenoides) dominated the interspaces (Hodgkinson 1983). At Boysag Point on the Grand Canyon’s North Rim, small-leaf globemallow occurred on swales (0.4% cover) and slopes (0.2% cover) but not on ridges. Soils on the swales and slopes were moderately developed calcareous, gravelly loams with lime accumulations at about 20 to 26 in (51-66 cm) deep. Soils on the ridges were undeveloped and reached depths of only about 6 in (15 cm) (Schmutz et al. 1967).
In an evaluation of vegetation and soils at Fishtail Mesa in the Grand Canyon, small-leaf globemallow occurred with Utah juniper (J. osteosperma), twoneedle pinyon (P. edulis), and big sagebrush (A. tridentata) in shallow (12-18 in [30-46 cm]), poorly developed, sandy to clay loam soils with pH levels of 8.2 to 8.7 (Jameson et al. 1962).
Description
Small-leaf globemallow is a perennial with a large taproot and a branching, stout, woody caudex (Fig. 2) (Kearney 1935; Welsh et al. 2015). Plants are sometimes referred to as subshrubs and produce several to many erect stems up to 3.3 ft (1 m) tall (Munz and Keck 1973; Holmgren et al. 2005; La Duke 2016). Stems are slender (≤ 5 mm in diameter at the base), unbranched or more commonly short-branched, and densely covered with stellate hairs, at least when young (Kearney 1935; Holmgren et al. 2005). Basal leaves are lacking; stems leaves are alternate (Munz and Keck 1973; Holmgren et al. 2005; Welsh et al. 2015). Larger leaves are 0.6 to 2.2 in (1.5-5.5 cm) long and about as wide with 0.4 to 2-in (1-5 cm) long petioles (Welsh et al. 2015). Leaf blades are thick and lack lobes or have three (sometimes five) shallow, broad, rounded lobes (La Duke 2016; LBJWC 2023). Leaves are covered with a stellate pubescence (La Duke 2016), prominently five-veined on the under sides, and regularly toothed margins (Fig. 3) (Kearney 1935).

Figure 2. Small-leaf globemallow growing in Utah. Photo: BLM UT040 SOS.

Figure 3. Small-leaf globemallow with visible pubescence on its stems growing in Utah. Photo: BLM UT391 SOS.

Figure 4. Small-leaf globemallow inflorescence with visible stamen columns on a plant growing in Utah’s Uintah Basin. Photo: S. Young, Utah State University (USU).
Flowers are numerous, clustered, and crowded in panicles (Fig. 4) (Munz and Keck 1973; LBJWC 2023). There are distinct internodes between the flower clusters and commonly more than one flower per node. Flower buds have a distinct beak (Kearney 1935; Welsh et al. 2015; La Duke 2016). The pedicel measures 2 to 10 mm long and is shorter than the densely pubescent calyx, which is 4 to 8 mm tall at anthesis (Kearney 1935; Munz and Keck 1973). Flowers have five orange to red petals each 7 to 18 mm long (Kearney 1935; Welsh et al. 2015; La Duke 2016). Flowers have many stamens. The filaments are united forming a column, which surrounds the pistils. The fruit is a segmented schizocarp (Fig. 5). The united stamen column and segmented schizocarps are characteristic of the Malvaceae (Weber 1976). Schizocarps are 4 to 5.5 mm in diameter and divided into 9 to 12 mericarps, each with two or sometimes one seed. Mericarps are ellipsoid, 3 to 5 mm high, and half or more as wide (Munz and Keck 1973; Holmgren et al. 2005; La Duke 2016). Seeds are kidney shaped, dark brown to black with a fine network of veins, 1.7 to 2 mm long, and more or less pubescent (Kearney 1935; Holmgren et al. 2005).

Figure 5. Small-leaf globemallow schizocarps dehiscing and leaving the mericarps visible in some cases and showing dispersal in others. These plants were growing at the Vermillion Cliffs area, near the Utah-Arizona border. Photo: S. Young, USU.
Because of hybridization, polyploidy, and morphological changes in response to environmental conditions (especially rainfall), globemallow species can be difficult to distinguish. Plants growing in Arizona are considered particularly difficult to identify (La Duke 2016). Small-leaf and Munro’s globemallow (S. munroana) are easily confused since they share many morphological characteristics, hybridize, and have overlapping distributions in Nevada, Utah, and Colorado. Small-leaf globemallow can be distinguished by its shallow leaf lobing (Sriladda et al. 2012). Although habitats can overlap, small-leaf globemallow often occurs at lower elevations and more southern locations than Munro’s globemallow (Welsh et al. 2015).
Reproduction
Small-leaf globemallow reproduces by seed. Its flowers are pollinated by insects, largely bees. Flowering is indeterminate and common from April through July (Ogle et al. 2011; LBJWC 2023).
Ecology
Small-leaf globemallow tolerates harsh conditions, disturbances, and grows rapidly (Ehleringer et al. 1991; Ogle et al. 2011; Huffman et al. 2013; Rink et al. 2020). In a study evaluating species growth and physiological survival strategies, small-leaf globemallow was classified as a suffrutescent subshrub with major summer dieback and near total dormancy during the summer drought between rainy spring and fall seasons (Comstock et al. 1988). Plants can green up if moisture interrupts dry weather (Pendery and Rumbaugh 1986). Small-leaf globemallow was monitored for 16 months beginning in April 1985 at two sites in western Arizona where it flowered in both the spring and fall. During this period, plants were not deciduous and produced considerable growth through the winter, which was unusually warm and mild. Plants maintained an above-ground woody caudex and had a robust growth form with stout, erect stems and relatively large leaves. Although most photosynthesis occurred in the leaves, the stems exhibited positive but low photosynthetic rates (Comstock et al. 1988). In a follow-up study assessing seasonal water use among different life forms, small-leaf globemallow competed directly with other life forms for limited water during drought conditions (Ehleringer et al. 1991). It utilized summer precipitation better than woody perennials in desert scrub vegetation in Utah’s Glen Canyon National Recreation Area. Researchers predicted that as summer precipitation made up a larger proportion of total precipitation, small-leaf globemallow may outcompete woody perennials (Ehleringer et al. 1991).
Although tolerant of harsh, dry, growing conditions, studies suggest that small-leaf globemallow abundance and seed production may be improved with higher moisture conditions (Pendery and Rumbaugh 1990; Beno 2009; Welsh et al. 2015). When plant growth was compared at two experimental pasture sites, one in southern Idaho and the other in northern Utah, small-leaf globemallow seed production was much greater at the Utah site that received more precipitation over the 4-year study (Pendery and Rumbaugh 1990). When vegetation composition was compared on cleared and untreated pinyon-juniper stands on a ranch in Seligman, Arizona, abundance of small-leaf globemallow was not associated with treatments and was greatest on the most level site. Frequency of small-leaf globemallow ranged from 0 to 1% on treated and untreated sites with slopes of 10 to 30%. Frequency was 28% at a recently logged oneseed juniper (Juniperus monosperma) site with almost no slope. The researcher suggested that reduced water runoff on the level site may have been most important to the high frequency of small-leaf globemallow (Beno 2009).
Seed And Seedling Ecology
Small-leaf globemallow emerged from soils collected from blackbrush/Indian ricegrass (Achnatherum hymenoides) vegetation in the Glen Canyon Recreation Area on the Utah-Arizona border. Soil samples were collected from early May to late June, suggesting that emergence was from soil-stored seed (Butler and Paintner 1991).
Disturbance Ecology
Globemallows are disturbance tolerant. When hazardous fuels treatment effects were evaluated for the understory of twoneedle pinyon-Utah juniper woodlands in northern Arizona’s Kaibab National Forest, globemallows were detected only after burning (3.3%) or burning and thinning (0.8%) operations. Globemallows remained at the treated sites for at least 5 years following treatments (Huffman et al. 2013).
While moderate disturbances are tolerated, pollution exposure may harm small-leaf globemallow. In experiments, plants suffered 20% damage after exposure to 0.5 ppm of sulfur dioxide, a common air pollutant from coal-fired power plants. Small-flower globemallow was one of the most sensitive species of the 87 native cold desert species evaluated (Hill et al. 1974).
Wildlife And Livestock Use
Globemallows are utilized by a variety of large and small mammals, birds, and reptiles (Gullion 1960; Hansen et al. 1976; Spaulding 1991; Beno 2009; Ogle et al. 2011). Small-leaf globemallow is considered good forage for sheep, goats, cattle, pronghorn, and other wildlife (Beno 2009; Cooney and Dreesen 2018). Based on pollen analysis of packrat middens (Neotoma spp.) in the Glen Canyon National Recreation Area in southeast Utah, Sphaeralcea species decreased from pre-settlement to post-settlement times when domestic grazers were introduced to area (Fisher et al. 2009).
Gambel’s quail (Callipepla gambelii) feed on globemallow seeds and herbage (Gullion 1960). In the Lower Grand Canyon, Arizona, globemallows made up 21% of the diets of desert tortoises (Gopherus agassizii) as determined from fecal analyses. Globemallows comprised 6% of the desert tortoise fecal samples collected from Arizona’s New Water Mountains but were not recovered from feces collected from Beaver Dam Wash in southwestern Utah (Hansen et al. 1976).
In the Harquahala Mountains of western Arizona, globemallows made up a large part of bighorn sheep (Ovis canadensis) and mule deer (Odocoileus hemionus) diets (Table 1). It was also a minor component (0.1-0.5%) of burro diets (Krausman et al. 1989).
Table 1. The percentage of bighorn sheep and mule deer diets comprised of globemallow species. Diets of male and female bighorn sheep were evaluated separately (Krausman et al. 1989).
|
Season |
Spring |
Summer |
Fall |
Winter |
|
———————diet %——————– |
||||
|
Bighorn sheep (male) Bighorn sheep (female) |
19.5 6.5 |
4.4 7.3 |
19.0 19.4 |
8.1 8.2 |
|
Mule deer |
9.4 |
6.7 |
24.0 |
13.2 |
Globemallow plant fragments made up a high proportion of the density of bighorn sheep feces (52.2%) but less of cattle (5.1%) and burro (4.1%) feces collected from the western end of the Lower Grand Canyon, Arizona (Hansen and Martin 1973).
The effects of grazing on small-leaf globemallow have been studied on a variety of sites in the West. In a new pasture near Kimberly, Idaho, sheep preferred alfalfa (Medicago sativa) over globemallows and globemallows over grasses in the spring but preferred alfalfa and grass over globemallows in the fall (Rumbaugh et al. 1993b). The pasture was transplanted with 3-month-old plants in April 1988, fall grazed in 1988 and 1989, and spring grazed in 1990 and 1991 by 10 to 15 ewes. Mortality of small-leaf globemallow plants between October 1988 and May 1989 averaged 57% for grazed plants and 40% for ungrazed plants. For the seven small-leaf globemallow accessions, mortality ranged from 36 to 75% for grazed and 0 to 75% for ungrazed plants (Rumbaugh et al. 1993b). Near the North Rim of the Grand Canyon, small-leaf globemallow was more abundant on less grazed sites when compared to heavily grazed sagebrush-dominated sites (Schmutz et al. 1967). The less-grazed site received some livestock grazing for about 20 years prior to the study and before that was grazed by about 40 sheep for 2 months per year. The heavily grazed site received livestock (sheep and cattle) and deer use continuously since the time of settlement. Cover of small-leaf globemallow was 0.2% on the largely ungrazed and 0.1% on the grazed site. Percent composition was 0.8% on the ungrazed and 0.3% on the grazed site (Schmutz et al. 1967).
Small-leaf globemallow is a good nectar and pollen plant for bees, especially globemallow bees (Diadasia diminuta) and is a host to the common checkered-skipper (Pyrgus communis) (Cooney and Dreesen 2018; Robinson et al. 2023). Vaudo et al. (2023) investigated pollen macronutrient content for a diversity of forb species to explain patterns of interactions among wild bee visitation and to aid in selection of of forbs to restore bee pollinator habitats. Sphaeralcea spp. had relatively low protein and moderate lipid levels which were associated with globemallow bees, sweat bees (Agapostemon spp.), small carpenter bees (Ceratina spp.), and furrow bees (Halictus spp.).
Nutritional Value
Globemallows are palatable to deer and livestock (Parkinson 2003), and several studies have evaluated their nutritional content. Average protein content of globemallows varies by season and site (Table 2; Krausman et al. 1989; Albert and Krausman 1993).
Table 2. Seasonal protein content (percent) of globemallows growing in the Harquahala (Krausman et al. 1989) and Belmont mountains (Albert and Krausman 1993) of Arizona.
|
Season |
Harquahala Mtns. |
Belmont Mtns. |
|
————protein content %————– |
||
|
Spring |
12.7 |
17.6 |
|
Summer |
15.7 |
12.4 |
|
Fall |
No data |
15.6 |
|
Winter |
16.7 |
11.5 |
Small-leaf globemallow growing with alfalfa or crested wheatgrass in new pastures near Kimberly, Idaho, met the nutrient requirements for domestic sheep and medium-sized, yearling heifers in the fall and spring (Table 3; Rumbaugh et al. 1993a).
Table 3. Nutritional content (evaluated and averaged for fall and spring) of small-leaf globemallow plants growing near Kimberley, Idaho (Rumbaugh et al. 1993a).
|
Measurement |
Microminerals (µg/g) |
Macrominerals (mg/g) |
||||||||
|
Element |
Cu |
Fe |
Mn |
Na |
Zn |
Ca |
Mg |
N |
P |
K |
|
Leaves |
15 |
1400 |
60 |
240 |
42 |
25 |
5.7 |
38 |
3.6 |
18 |
|
Stems |
16 |
440 |
23 |
470 |
32 |
14 |
5.4 |
22 |
2.3 |
22 |
Ethnobotany
The Havasupai and Hopi people used small-leaf globemallow as a color additive, medicine, and in ceremonies (Moerman 2003). Weber and Seaman (1985) indicated that the Havasupai mixed clay with juice of the plant before molding it into a pot. Colton (1974) noted that the Hopi chewed the roots or boiled them with cactus root to help ease constipation in adults and babies. Small-leaf globemallow was used to treat sores, cuts, wounds, and broken bones. It was also used in mid-winter ceremonies (Colton 1974).
Horticulture
Small-leaf globemallow is drought tolerant with a long bloom period, making it a good choice for use in low-water landscapes (Cooney and Dreesen 2018). At a native plant garden in Boise, Idaho, globemallow plants grew well when provided supplemental water four times each month in the establishment year and once a month in subsequent years. Plants that were cut back after flowering were compact and bloomed prolifically (Parkinson 2003).
Revegetation Use
As a colonizer of disturbed sites, small-leaf globemallow is used for erosion control in semi-arid and arid lands in the Southwest. Small-leaf globemallow has a rapid growth rate, provides a nectar and pollen source for pollinators, and is palatable to livestock and wildlife, especially sheep, goats, and pronghorn (Cooney and Dreesen 2018). Small-leaf globemallow seed is commercially available (Dorn and Dorn 2007) and it is adapted to restoration sites in the 6 to 15 in (152-381 mm) annual precipitation zone (Tilley et al. 2013; Ogle et al. 2019).
Developing A Seed Supply
Substantial amounts of small-leaf globemallow seed can be harvested from native stands (S. Young, Utah State University [retired], personal communication, August 2018). Though production is sporadic, seed has been harvested using mechanical methods on extensive stands south of the Vermillion Cliffs in northern Arizona and in the south-central part of Utah’s Uintah Basin (Fig. 6).
In a study of the spatial and landscape genetics of native plant species in Arches and Canyonlands National Parks, research was conducted on 22 small-leaf globemallow populations. Total genetic variation was 84% within and 16% among small-leaf globemallow populations, which was significantly different from the random expectation (P < 0.001). The association between genetic and geographic distances was poor, suggesting a high level of gene flow among populations separated by relatively large geographical areas through pollinator-mediated, long-distance pollen transfer (Choo 2012).

Figure 6. Extensive stand of small-leaf globemallow growing in the south-central portion of Utah’s Uintah Basin. Photo: S. Young, USU.
For restoration to be successful, the right seed needs to be planted in the right place at the right time. Coordinated planning and cooperation is required among partners to first select appropriate species and seed sources and then properly collect, grow, certify, clean, store, and distribute seed for restoration (PCA 2015).
Developing a seed supply begins with seed collection from native stands. Collection sites are determined by current or projected revegetation requirements and goals. Production of nursery stock requires less seed than large-scale seeding operations, which may require establishment of agricultural seed production fields. Regardless of the size and complexity of any revegetation effort, seed certification is essential for tracking seed origin from collection through use (UCIA 2015).
Seed Sourcing
Massatti (2019) provides genetically-based seed transfer zones and accompanying environmental variables influencing adaptation across most of small-leaf globemallow’s distribution. For regions outside of this range, generalized provisional seed zones developed by Bower et al. (2014), may be used to select and deploy seed sources. These provisional seed zones identify areas of climatic similarity with comparable winter minimum temperature and aridity (annual heat:moisture index). In Figure 7, Omernik Level III Ecoregions (Omernik 1987) overlay the provisional seeds zones to identify climatically similar but ecologically different areas. For site-specific disturbance regimes and restoration objectives, seed collection locations within a seed zone and ecoregion may be further limited by elevation, soil type, or other factors.
The Western Wildland Environmental Threat Assessment Center’s (USFS WWETAC 2024) Threat and Resource Mapping (TRM) Seed Zone application provides links to interactive mapping features useful for seed collection and deployment planning. The Climate Smart Restoration Tool (Richardson et al. 2020) can also guide restoration planning, seed collection, and seed deployment, particularly when addressing climate change considerations.
In transplanted pastures in northern Utah and southern Idaho, survival and growth of 17 small-leaf globemallow accessions and three small-leaf × Munro’s globemallow hybrids were compared. Average transplant survival to about 1.5 years for all small-leaf globemallow accessions was high (92%), but was only 71% for one accession from seed collected from southern Utah. Researchers suggested that seed from this southern location was poorly adapted to more northern conditions (Pendery and Rumbaugh 1990).
Occurrence Map
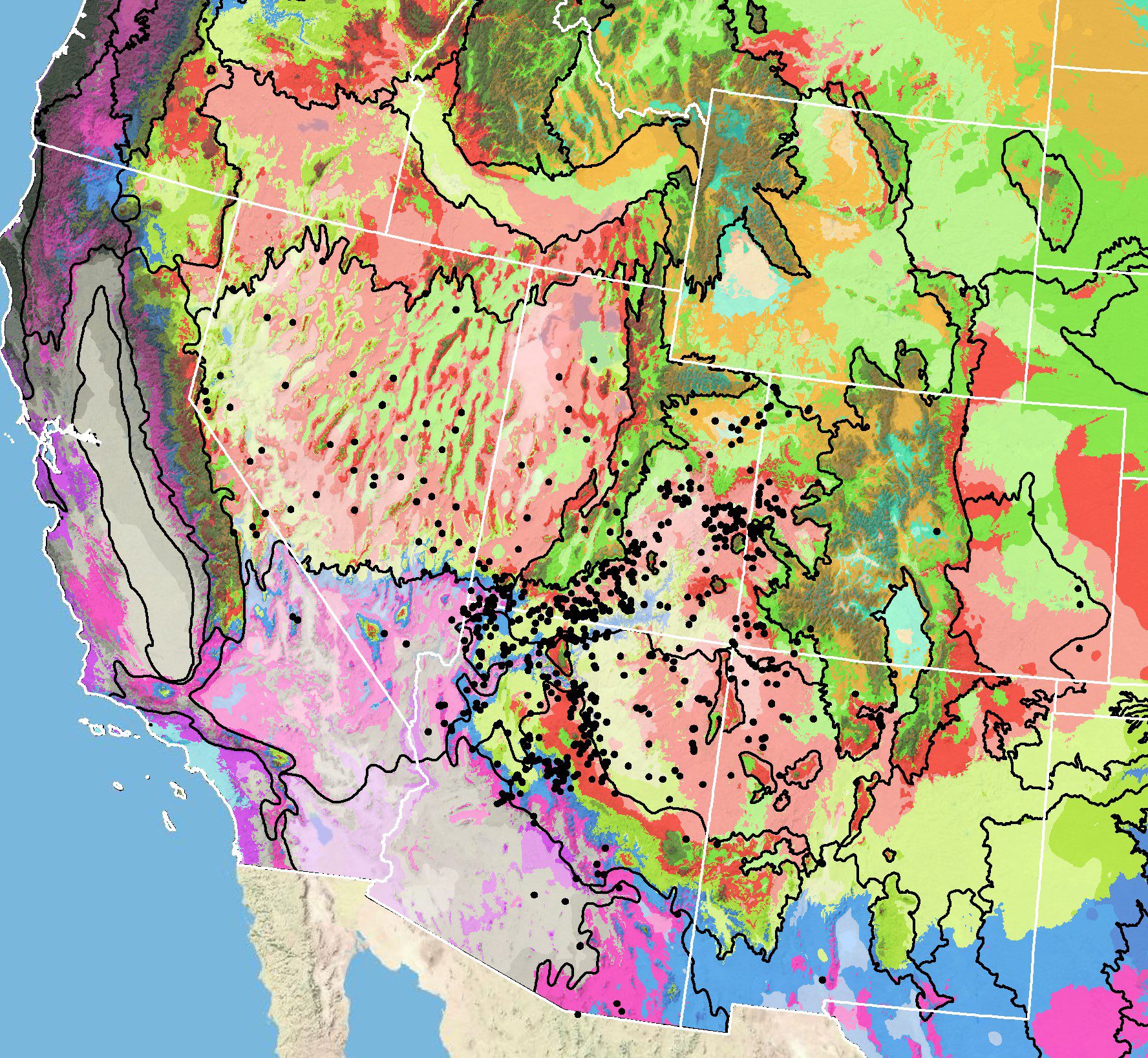
Figure 7. Distribution of small-leaf globemallow (black circles) based on geo-referenced herbarium specimens and observational data from 1911-2016 (CPNWH 2017; SEINet 2017; USGS 2017). Generalized provisional seed zones (colored regions) (Bower et al. 2014) are overlain by Omernik Level III Ecoregions (black outlines) (Omernik 1987; USDI EPA 2018). Interactive maps, legends, and a mobile app are available (USFS WWETAC 2024; research.fs.usda.gov/pnw/products/dataandtools/seed-zone-webmap). Map prepared by M. Fisk, USGS.
Releases
As of 2025, there were no releases of small-leaf globemallow.
Wildland Seed Collection
Small-leaf globemallow flowers indeterminately with flowers lower on the inflorescence opening before the upper flowers. This creates a wide harvest window at some locations (Fig. 5). Mechanical harvests are possible in dense native stands (Fig. 8). Dense stands growing near the Utah-Arizona border have been harvested mechanically using strippers with hoppers (Figs. 9, 10). In less dense or less easily accessible stands, wildland seed can be hand collected by clipping or plucking ripe schizocarps (Fig. 5).
Wildland Seed Certification
Verification of species and tracking of geographic source is necessary whether wildland seed is collected for immediate project use or as stock seed for cultivated increase. This official Source Identification process can be accomplished by following procedures established by the Association of Official Seed Certifying Agencies (AOSCA) Pre-Variety Germplasm Program (UCIA 2015; Young et al. 2020). Wildland seed collectors should become acquainted with state certification agency procedures, regulations, and deadlines in the states where they collect.
If wildland-collected seed is to be sold for direct use in ecological restoration projects, collectors must apply for Source-Identified certification prior to making collections. Pre-collection applications, site inspections, and species and seed amount verification are handled by the AOSCA member state agency where seed collections will be made (see listings at AOSCA.org).
If wildland seed collected by a grower or private collector is to be used as stock seed for planting cultivated seed fields or for nursery propagation (See Agricultural Seed Field Certification section), detailed information regarding collection site and collecting procedures must be provided when applying for certification. Photos and herbarium specimens may be required. Germplasm accessions acquired within established protocols of recognized public agencies, however, are normally eligible to enter the certification process as stock seed without routine certification agency site inspections. For contract grow-outs, however, this collection site information must be provided to the grower to enable certification.
Collection Timing
Collections can be maximized by harvesting the schizocarps when the lowest ones have started to split and most schizocarps are just starting to open. At this time, schizocarps will be light green to brown and about 25% of the seeds will be mature (Pendery and Rumbaugh 1993). Because seeds disperse once schizocarps are fully mature, populations should be monitored closely once plants begin to flower (Shock et al. 2008). Seed harvests may also be impacted by weevils (Apionidae), some of which are specialist globemallow seed predators. They can severely limit seed production and their populations should be monitored when inspecting stands for potential harvest (CSUE 2016).
The BLM SOS program made 69 collections of small-leaf globemallow seed between 2003 and 2024 (USDI BLM SOS 2017; 2024). Most collections were made in Utah (52) but others were made in Arizona (9), Nevada (7), and Colorado (1). Although collection dates ranged from late May to early November, most collections were made in June (34) and July (27). At several sites collectors harvested more than once, sometimes days or up to 3 weeks following the first collection. The earliest collection date was May 26, 2015, made in Grand County, Utah, at 4,588 ft (1,398 m) elevation and the latest collection date was November 4, 2018, made in Arizona (county not recorded) at 4,801 ft (1,463 m) elevation. In the years with the most collections made, 2010 and 2015, each with 10 harvests, the earliest collection date and location in 2010 was June 22 made in Uintah County, Utah, at 5,074 ft (1,547 m) elevation and the latest in 2010 was on August 11 in Coconino County, Arizona, at 6,317 ft (1,925 m). In 2015, earliest harvest date and location was May 26, 2015, made in Grand County, Utah at 4,588 ft (1,398 m) elevation and the latest was July 15 in Kane County, Utah, at 5,057 ft (1,541 m) (USDI BLM SOS 2017; 2024).
In small-leaf globemallow stands established from transplants, flowering dates varied by site and plant age (Pendery and Rumbaugh 1990). First flowering was earlier at a northern Utah (early June) than at a southern Idaho site (late June). Average date of first flowering at both sites was 1.5 months earlier (June 1) for second year plants than it was for plants in their establishment year (July 15). Researchers also reported drier conditions in the second year (Pendery and Rumbaugh 1990).

Figure 8. Dense wildland stand of small-leaf globemallow setting seed at the Vermillion Cliffs area near the Utah-Arizona border. Photo: S. Young, USU.

Figure 9. Strippers with hoppers used to mechanically harvest dense small-leaf globemallow stands growing near the Utah-Arizona border. Photo: S. Young, USU.
Weather patterns in western Arizona may allow for spring and fall seed harvests (Comstock et al. 1988). When small-leaf globemallow was monitored at two Mojave Desert sites, plants flowered in both the spring and fall, which coincided with the bimodal rainy periods. Flowering peaked in April. Plants in this area experienced considerable dieback and were essentially dormant through the summer drought conditions. The winter during this one-year study period was also uncharacteristically warm and mild (Comstock et al. 1988).

Figure 10. Mechanically harvested small-leaf globemallow seed from dense stands growing near the Utah-Arizona border. Photo: S. Young, USU.
Collection Methods
Several collection guidelines and methods should be followed to maximize the genetic diversity of wildland collections: 1) collect seed from a minimum of 50 randomly selected plants; 2) collect from widely separated individuals throughout a population without favoring the most robust or avoiding small stature plants; and 3) collect from all microsites including habitat edges (Basey et al. 2015). General collecting recommendations and guidelines are provided in online manuals (e.g., ENSCONET 2009; USDI BLM SOS 2023).
It is critical that wildland seed collection does not impact the sustainability of native plant populations. Collectors should take no more than 20% of the viable seed available at the time of harvest (USDI BLM SOS 2023). Additionally, care must be taken to avoid the inadvertent collection of weedy species, particularly those that produce seeds similar in shape and size to those of small-leaf globemallow.
Post-Collection Management
Seed should be kept in a dry, shaded place until collections can be moved to a controlled short-term storage environment, where seed collections should be dried fully (Gold n.d.; Hay and Probert 2011). If collections include plant material, which greens up following summer precipitation, more ventilation and drying time will be required. Because the seed is attractive to weevils (Macrophoptus spp.), it should be inspected prior to storage and treated or frozen to suspend further insect damage. Storage areas should be protected from insects and small mammals (Pendery and Rumbaugh 1986).
Seed Cleaning
Dry seed can be cleaned using a debearder to remove seeds from the schizocarps. If necessary, a clipper or fanning mill can be used to further clean the seed (Pendery and Rumbaugh 1993). Cleaned seed are shown in Figure 11.
At the Los Lunas Plant Materials Center (LLPMC), a small-leaf globemallow seed production plot was harvested using a forage harvester (Cooney and Dreesen 2018). Harvested material which was still green was dried then processed through a hammermill using screen size of 0.25 in at a dial speed of 8. This process dislodged seed from the carpels and reduced the volume of plant mass. Remaining plant material and seeds were then processed to pure live seed (PLS) quality using a four-screen fanning mill using 17, 12, 1/17, and 1/25 screen sizes (Cooney and Dreesen 2018).
Seed Storage
Small-leaf globemallow seed is orthodox. Seed viability was 95% after seeds were dried and stored at -4 °F (-20 °C) for about 2 years (SER, INSR, RBGK 2023).
Seed Testing
There is no AOSA germination protocol for small-leaf globemallow (AOSA 2016), but AOSA guidelines for tetrazolium chloride (TZ) viability testing of the Sphaeralcea genus are available. Seed is prepared by soaking it overnight at 68 to 77 °F (20-25 °C) followed by staining with 0.1 or 1% TZ. Seed is nonviable if any part of the embryo or endosperm is unstained. Clipping prior to soaking and staining is recommended for hard seeds (AOSA 2010).

Figure 11. Clean small-leaf globemallow seed. Photo: BLM CP2 SOS.
Germination Biology
Scarification is generally necessary to obtain rapid and high levels of small-leaf globemallow germination. Low temperature exposure following scarification may improve germination of seed collected from more northern locations. High levels of germination (95%) were reported after 28 days when a small seed sample was scarified and then incubated at 59 °F (15 ºC) or 68 °F (20 °C) with an 8 hr/16 hr light/dark cycle. Seed scarification involved partial removal or chipping of the seed coat with a scalpel (SER, INSR, RBGK 2023). Based on germination studies conducted by Norman C. Deno, germination of small-leaf globemallow seed was 100% after 2 days at 70 °F (21 °C) when a hole was ground through the seed coat. Germination was 4% at 40 (4 °C) or 70 °F (21 °C) when the seed coat was not penetrated (Love and Akins 2020).
Scarification followed by cold temperature exposure (34 °F [1 °C] for ≥4 wks) resulted in the greatest germination (≥ 70%) of small-leaf globemallow seed collected from Uinta County, Utah. Unscarified seeds subjected to extended periods of cold (12 weeks) had lower germination percentages (≤ 35%), and scarified seed exposed to warmer temperatures (≥41 °F [5 °C]) for 12 weeks also had lower germination percentages (≤ 30%). The scarification method was not described (Kramer and Foxx 2016).
Germination of small-leaf globemallow was improved by mechanical scarification; stratification treatments provided no additional improvement (Table 4; Seglias et al. 2018). The study was conducted to identify relationships between source climate, species identity, and germination to improve seed sourcing decisions in restoration; small-leaf globemallow was one of several species evaluated. Seed came from eastern Utah where annual temperature averages 48 °F (8.9 °C) and precipitation averages 18 in (465 mm) and from near Flagstaff, Arizona, where annual average temperature was similar (54 °F [12 °C]) but precipitation was less (9 in [229 mm]). Seed was afterrippened for 2 weeks, dried, scarified with sandpaper (15-30 s), then exposed to various stratification treatments mimicking long winter, spring germination; short winter, spring germination; and short summer monsoon and fall germination conditions followed by spring, summer, or fall germination temperatures (Table 4; Seglias et al. 2018).
Researchers at the Great Basin Research Center reported that no pre-treatments were necessary to germinate small-leaf globemallow seed. Collection location, seed storage conditions (if any), and germination conditions were not reported (Vernon et al. 2006).
Table 4. Germination of afterripened (2 wks) and mechanically scarified seeds exposed to selected stratification and incubation conditions (4 replicates of 25 seeds per treatment). No statistical analysis was conducted (Seglias et al. 2018).
|
Simulated conditions |
Control |
Long winter, spring germ |
Short winter, spring germ |
Short summer monsoon, fall germ |
||||
|
Stratification (time, temp): |
none |
8 wks, 37 °F |
3 wks, 37 °F |
3 wks, 86 °F |
||||
|
Incubation*: |
cool |
warm |
cool |
warm |
cool |
warm |
cool |
warm |
|
Seed source |
———————————————-germination %—————————————— |
|||||||
|
Flagstaff, AZ |
75 |
79 |
38 |
36 |
38 |
59 |
68 |
71 |
|
Eastern UT |
86 |
85 |
74 |
80 |
47 |
48 |
44 |
64 |
*Cool (spring/fall) incubation was 68/50 °F (20/10 °C) and warm (summer) was 77/59 °F (25/15 °C) with 12 hr/12 hr dark/light cycle.
Wildland Seed Yield And Quality
Post-cleaning seed yield and quality of small-leaf globemallow seed lots collected in the Intermountain region are provided in Table 5 (USFS BSE 2017, 2023). The results indicate that small-leaf globemallow can generally be cleaned to high levels of purity and seed fill but that viability of fresh seed is variable. Other sources (Ogle et al. 2011; USFS GBNPP 2014; SER INSR RBGK 2023) reported that small-leaf globemallow seed weights fell within the range reported in Table 5.
Table 5. Seed yield and quality of small-leaf globemallow seed lots collected in the Intermountain region, cleaned and evaluated by the Bend Seed Extractory. Viability was tested by the Oregon State Seed Laboratory or the USFS National Seed Laboratory (USFS BSE 2017, 2023).
|
Seed lot characteristic |
Mean |
Range |
Samples (no.) |
|
Bulk weight (lbs) |
0.99 |
0.04-3.34 |
68 |
|
Clean weight (lbs) |
0.14 |
0.0005-0.65 |
68 |
|
Clean-out ratio |
0.13 |
0.0014-0.27 |
68 |
|
Purity (%) |
98 |
87-99 |
68 |
|
Fill (%)¹ |
89 |
5-99 |
68 |
|
Viability (%)² |
92 |
44-100 |
61 |
|
Seeds/lb |
405,701 |
189,997-630,000 |
68 |
|
Pure live seeds/lb |
360,270 |
176,811-550,323 |
61 |
¹100 seed X-ray test
²Tetrazolium chloride test
Marketing Standards
Acceptable seed purity, viability, and germination specifications vary with revegetation plans. Purity needs are highest for precision seeding equipment used in nurseries, while some rangeland seeding equipment handles less clean seed quite well. In 2007, the standardized market price for small-leaf globe mallow was $60/lb PLS (Young and Bouck 2007).
Agricultural Seed Production
Small-leaf globemallow seed production fields were grown successfully at the Oregon State University’s Malheur Experiment Station (OSU MES). Harvestable crops were produced for 5 years. Irrigation increased seed yields but not significantly. Although annual flailing of fields in the fall concentrated the seed production period for a single annual harvest, multiple hand harvests resulted in greater yields than a single mechanical harvest (Shock et al. 2015).
The Los Lunas Plant Materials Center (LLPMC) established a dense 0.5 ac (0.2 ha) seed production stand in 2012 using greenhouse-grown small-leaf globemallow seedlings, which were planted into rows tilled 18 in (46 cm) deep using a straight shank subsoiler. Weeds were controlled by cultivation and hand-hoeing. Stands were irrigated weekly from August through September and then biweekly through November (Dreesen 2016 cited in Cooney and Dreesen 2018).
Agricultural Seed Certification
In order to minimize genetic changes in specific accessions of native species when increased in cultivated fields, it is essential to track the geographic source and prevent inadvertent hybridization or selection pressure. This is accomplished by following third party seed certification protocols for Pre-Variety Germplasm (PVG) as established by the Association of Official Seed Certification Agencies (AOSCA). AOSCA members in the U.S., Canada, and other countries administer PVG requirements and standards that track the source and generation of planting stock. Field and cleaning facility inspections then monitor stand establishment, proper isolation distances, control of prohibited weeds, seed harvesting, cleaning, sampling, testing, and labeling for commercial sales (UCIA 2015; Young et al. 2020).
Seed growers apply for certification of their production fields prior to planting and plant only certified stock seed of an allowed generation (usually less than four). The systematic and sequential tracking through the certification process requires preplanning, knowing state regulations and deadlines, and is most smoothly navigated by working closely with state certification agency personnel. See the Wildland Seed Certification section for more information on stock seed sourcing.
Weed Management
At OSU MES, weeds in seed production plots were controlled by hand weeding (Shock et al. 2015). In a study evaluating rehabilitation of cheatgrass (Bromus tectorum)-invaded shrublands, findings suggest that small-leaf globemallow may be sensitive to imazapic herbicides (Owen et al. 2011). Small-leaf globemallow was a dominant species in the mixed shrub community on the Kaibab Plateau in northern Arizona burned about 11 years prior to initiation of the study. Rehabilitation treatments included: a fall herbicide treatment with imazapic, drill seeding with big sagebrush and Mexican cliffrose (Purshia mexicana), a combined treatment, and a control. An evaluation of the nontarget effects revealed average cover of small-leaf globemallow was half as much on herbicide and herbicide-seeded plots as on untreated and seeded plots (P < 0.0001) and was reduced for at least 2 years (Owen et al. 2011).
Seeding
Fall seeding is recommended for small-leaf globemallow. Seeding in the spring (April) resulted in poor stand emergence. Spring sowing was done using non-scarified seed soaked in distilled water, rinsed in bleach solution, and stratified for about 2.5 months. Sowing untreated seed in the fall resulted in good small-leaf globemallow stands (Shock et al. 2008, 2015). At OSU-MES, 30 seeds were planted per foot of row in weed-free fields (Shock et al. 2015).
Irrigation
Based on subsurface drip irrigation trials, researchers concluded that small-leaf globemallow needed less than 4 in (102 mm) of supplemental irrigation per year when growing at OSU MES in Ontario, Oregon (Shock and Feibert 2016). Seed yield of seed production fields at OSU MES increased, but not significantly, with supplemental annual irrigation beyond water provided during the establishment period (Table 6; Shock et al. 2015). Irrigation research findings suggested that natural rainfall in Ontario, Oregon, allowed for maximum seed production in weed-free conditions (Table 7).
Table 6. Seed yield (lbs/ac) for fall-seeded (November 2006) small-leaf globemallow in response to no or 0, 4, or 8 in irrigation at Oregon State University’s Malheur Experiment Station in Ontario, OR (Shock et al. 2015).
|
Year |
Supplemental irrigation (in/season*) |
||
|
0 |
4 |
8 |
|
|
——————yield (lbs/ac)—————– |
|||
|
2007** |
1062.6 |
850.7 |
957.9 |
|
2008 |
436.2 |
569.1 |
554.7 |
|
2009 |
285.9 |
406.1 |
433.3 |
|
2010 |
245.3 |
327.3 |
257.9 |
|
2011 |
81.6 |
142.5 |
141.2 |
*Irrigation season was from floral bud to seed set and water was delivered through a subsurface drip irrigation system.
**2007 data represents the total of three separate hand harvests. Data for 2008-2011 represent single combine harvests.
Table 7. Weather conditions at Oregon State University’s Malheur Experiment Station in Ontario, OR (Shock et al. 2015).
|
Year |
Precipitation (in) |
Growing-degree hrs (50-86 °F) |
|
|
Jan-Jun |
Apr-Jun |
Jan-Jun |
|
|
2006 |
9.0 |
3.1 |
1120 |
|
2007 |
3.1 |
1.9 |
1208 |
|
2008 |
2.9 |
1.2 |
936 |
|
2009 |
5.8 |
3.9 |
1028 |
|
2010 |
8.3 |
4.3 |
779 |
|
2011 |
8.3 |
3.9 |
671 |
|
Mean |
5.8 |
2.7 |
1042 |
Pollinator Management
Small-leaf globemallow is pollinated by bees. Globemallow bees (Diadasia diminuta) are very effective pollinators and will eventually colonize and multiply in globemallow seed production fields (Cane personal communication cited in (Cooney and Dreesen 2018). To enhance ground-nesting habitat of globemallow bees, cultivations should be limited in and around seed fields.
Pest Management
Small-leaf globemallow is host to several pests that can reduce plant growth and seed production. Yet, some weevils (Apionidae) are specialist globemallow seed predators and can severely limit seed production (CSUE 2016). At OSU MES, a rust fungus, P. sherardiana, infected stands and caused substantial leaf loss and reduced vegetative growth in plants for 2 of the 5 years of production (Shock et al. 2015). Other fungal pathogens that utilize globemallow hosts include: Puccinia sherardiana, P. sphaeralceoides, P. platyspora, P. interveniens, Synchytrium spp., and Sphaerella stenospora (Farr and Rossman 2017). Globemallow leaf beetles (Calligrapha serpentina) feed on Sphaeralcea spp. leaves and lay eggs on the undersides of leaves, but leaf beetle impacts on seed production were not described (Cooney and Dreesen 2018).
Seed Harvesting
In an effort to maximize seed harvests, researchers evaluated seed yield and germination of immature to mature schizocarps in seed production plots in Utah. Immature schizocarps yielded the smallest percentage of ripe seed (3%), but this seed had the highest germination (50%). More mature, but not fully mature, schizocarps yielded 40% mature seed, which germinated at 28%. Fully mature schizocarps yielded the highest amounts of mature seed (45%) but had the lowest germination (12%). Seeds from the least mature schizocarps may have had thinner seed coats or less well-developed seed dormancy. More mature schizocarps may have required treatments not applied in this germination experiment (Meyer and Vernon 2008).
Seed fields grown at OSU MES were combine harvested (Shock et al. 2015) and at LLPMC they were harvested using a forage harvester (Cooney and Dreesen 2018).
Seed Yields And Stand Life
For small-leaf globemallow seed production stands growing at OSU MES, seed yield was greatest in the establishment year following fall seeding, but this was primarily the result of multiple hand seed collections. In following years, crops were flailed each fall to concentrate the timing of flowering, reduce duration of flowering, and allow for a single mechanical harvest with a combine in the following year (Table 8; Shock et al. 2015). Plants produced harvestable seed for 5 years (Shock et al. 2012, 2015), but by the fifth year seed production was greatly reduced. In the sixth year, growth was poor and the stands were eliminated (Shock et al. 2015).
Table 8. Flowering dates, harvest dates, and seed yields for seed production fields provided no supplemental irritation at Oregon State University’s Malheur Experiment Station in Ontario, OR (Shock et al. 2015).
|
Year |
Start of flowering |
End of flowering |
Harvest date(s) |
Seed yield (lbs/ac) |
|
2007 |
5 May |
30 Sept |
20 June, 10 July, 13 Aug |
1062.6* |
|
2008 |
5 May |
15 June |
21 July |
436.2 |
|
2009 |
1 May |
10 June |
14 July |
285.9 |
|
2010 |
10 May |
25 June |
20 July |
245.3 |
|
2011 |
26 May |
14 July |
29 July |
81.6 |
*Seed yield represents multiple hand-harvests, which is likely the reason for high yields and not year, plant age, or other factors.
Nursery Practice
Small-leaf globemallow has been grown successfully in the greenhouse (Pendery and Rumbaugh 1990; Vernon et al. 2005). Vernon et al. (2005) reported excellent emergence after direct seeding small-leaf globemallow into flats in the greenhouse. Scarified seed was planted 0.25 to 0.5 in (0.6-1.3 cm) deep. Pendery and Rumbaugh (1990) planted sandpaper-scarified seed in 9 in³ (150 cm³) cones filled with four parts sand, two parts peat moss, and one part vermiculite to produce seedlings for research studies.
Small-leaf globemallow seedlings were grown by the LLPMC from scarified seed (Cooney and Dreesen 2018) using an MAT-OSU pneumatic scarifier (Seedburo, Des Plaines, IL) for 60 s at 20 psi. Scarified seed was sown into moist soil in plug trays. About a half teaspoon of seed was mixed with about 4 teaspoons of grit, and this mixture was sieved evenly over each tray. Trays were covered with clear plastic and cold stratified at 39 °F (4 °C) for about 6 weeks before being moved to a greenhouse kept at 70 °F (21 °C) during the day and 55 °F (13 °C) at night. Once root masses were sufficient, trays were transferred to a shade house to harden off before they were transplanted in the field (Cooney and Dreesen 2018).
Wildland Seeding And Planting
Small-leaf globemallow is a recommended restoration species within the 6 to 15 in (152-381 mm) annual precipitation zone because of its rapid growth rate, value as forage, and importance to bees and other pollinators (Tilley et al. 2013). Fall seeding in medium to coarse soils is recommended (Monsen and Stevens 2004; Ogle et al. 2011). Seed can be broadcast, drill, or compact surface seeded as part of a seeding mixture or interseeded in existing vegetation provided enough area is cleared. Small-leaf globemallow is compatible with most other seeded species, and it has moderate seedling vigor (Monsen and Stevens 2004).
Monsen and Stevens (2004) recommend planting small-leaf globemallow seed 0.12 to 0.25-in (0.3-0.6 cm) deep in rows spaced 15 to 18 in (38-46 cm) apart. Ogle et al. (2011) suggested slightly deeper seeding (0.25-0.5 in [0.6-1.3 cm]) and wider row spacing (24-48 in [61-122 cm]). The recommended pure stand seeding rate is 2 lbs PLS/ac (2.25 kg/ha) (Ogle et al. 2011).
Use of small-leaf globemallow and resulting establishment was limited in the reviewed literature, but it is worth noting that small-leaf globemallow specimens collected in Cache County, Utah, in 1993 and in Oneida County, Idaho, in 2021 are outside of the native range of the species as they are persistent populations from experimental restoration plantings made by Pendery and Rumbaugh (1990) near these locations in 1987 (Daines, Anzjon et al. 2022).
Specimens collected near Curlew National Grassland in Oneida Co, ID in 2021. Reportedly planted experimentally by Pendery and Rumbaugh 1990 at this location in 1987. This location as well as Cache Co, UT collection made in 1993 represent persistent populations from restoration plantings (Daines, Anzjon et al. 2022)
Transplant survival of small-leaf globemallow was high (92%) a year after planting in dryland pastures planted for livestock grazing studies in southern Idaho and northern Utah (Pendery and Rumbaugh 1990). After 3 months of growth in the greenhouse, seedlings were planted into clean-tilled sites in mid-April 1987. In summer 1988, small-leaf globemallow transplants averaged 114 g (range: 82-139 g), 17 stems (range: 10-23), and had an average maximum stem length of 19 in (47 cm) (range: 5-22 in [12-55 cm]). Plant biomass was greater, plants flowered earlier, and seed production was greater at the Utah site where annual precipitation was 2 to 16 in (59-405 mm) greater between 1985 and 1988 (Pendery and Rumbaugh 1990).
Acknowledgements
Funding for Western Forbs: Biology, Ecology, and Use in Restoration was provided by the USDI BLM Great Basin Native Plant Materials Ecoregional Program through the Great Basin Fire Science Exchange. Great thanks to the chapter reviewers: Wayne Padgett, BLM (retired) and Stanford Young, Utah State University (retired).
This research was supported in part by the USDA Forest Service, Rocky Mountain Research Station. The findings and conclusions in this publication are those of the authors and should not be construed to represent any official USDA or U.S. Government determination or policy.
Literature Cited
Albert, S.K.; Krausman, P.R. 1993. Desert mule deer and forage resources in southwest Arizona. The Southwestern Naturalist. 38(3): 198-205.
Association of Official Seed Analysts [AOSA]. 2010. AOSA/SCST Tetrazolium testing handbook. Contribution No. 29. Lincoln, NE: Association of Official Seed Analysts.
Association of Official Seed Analysts [AOSA]. 2016. AOSA rules for testing seeds. Vol. 1. Principles and procedures. Washington, DC: Association of Official Seed Analysts.
Basey, A.C.; Fant, J.B.; Kramer, A.T. 2015. Producing native plant materials for restoration: 10 rules to collect and maintain genetic diversity. Native Plants Journal. 16(1): 37-53.
Beatley, J.C. 1975. Climates and vegetation pattern across the Mojave/Great Basin desert transition of southern Nevada. American Midland Naturalist. 93(1): 53-70.
Beno, A.L. 2009. A comparative study of rangeland browse and forage within a pinyon-juniper ecosystem in Seligman, Arizona. Prescott, AZ: Prescott College. Thesis. 49 p.
Bower, A.D.; St. Clair, J.B.; Erickson, V. 2014. Generalized provisional seed zones for native plants. Ecological Applications. 24(5): 913-919.
Butler, J.L.; Paintner, K.J. 1991. Rangeland recovery potential: Soil seed content and seed viability. In: Plumb, G.E., ed. University of Wyoming: National Park Service Research Center 15th annual report–1991. Laramie, WY: University of Wyoming: 130-135.
Cane, J.H. 2008. Pollinating bees crucial to farming wildflower seed for U.S. habitat restoration. In: James, R.; Pitts-Singer, T., eds. Bees in agricultural ecosystems. Oxford, UK: Oxford University Press: 48-64.
Choo, M. 2012. Spatial/landscape genetics of native plant species in Arches and Canyonlands National Parks. Memphis, TN: University of Memphis. 55 p.
Clark, A.; Burgess, R.L. 1966. The persistent perennial vegetation on the rim of Montezuma Well, Arizona. Journal of the Arizona Academy of Science. 4(1): 35-42.
Colorado State University Extension [CSU Ext.]. 2016. Western Colorado insects: Native plant seed production. http://www.wci.colostate.edu/seed_production.shtml
Colton, H.S. 1974. Hopi history and ethnobotany. In: Horr, D.A., ed. Hopi Indians. New York, NY: Garland Publishing: 279-386.
Comstock, J.P.; Cooper, T.A.; Ehleringer, J.R. 1988. Seasonal patterns of canopy development and carbon gain in nineteen warm desert shrub species. Oecologia. 75(3): 327-335.
Consortium of Pacific Northwest Herbaria [CPNWH]. 2017. Seattle, WA: University of Washington Herbarium, Burke Museum of Natural History and Culture. pnwherbaria.org/
Daines, M.; Anzjon, A.; Gay, M.; Headings, L.; Huckabee, J.; Marine, M.; Poolman, T.; Scoggins, S.; Styers, R.; Villafane, I.; Snow, N. 2022. Numerous distributional novelties collected by students for Idaho, Kansas, Missouri, Oklahoma, and Texas, U.S.A. Journal of the Botanical Research Institute of Texas. 16(2): 551-557.
Ehleringer, J.R.; Phillips, S.L.; Schuster, W.S.F.; Sandquist, D.R. 1991. Differential utilization of summer rains by desert plants. Oecologia. 88(3): 430-434.
European Native Seed Conservation Network [ENSCONET]. 2009. ENSCONET seed collecting manual for wild species. Edition 1: 32 p.
Farr, D.F.; Rossman, A.Y. 2017. Fungal databases, U.S. National Fungus Collections. U.S. Department of Agriculture, Agricultural Research Service. https://nt.ars-grin.gov/fungaldatabases/.
Fisher, J.; Cole, K.L.; Anderson, R.S. 2009. Using packrat middens to assess grazing effects on vegetation change. Journal of Arid Environments. 73(10): 937-948.
Gold, K. n.d. Post-harvest handling of seed collections. Technical Information Sheet 04. UK: Royal Botanic Gardens Kew and Millennium Seed Bank Partnership. 4 p.
Gullion, G.W. 1960. The ecology of Gambel’s quail in Nevada and the arid southwest. Ecology. 41(3): 518-536.
Hansen, R.M.; Johnson, M.K.; Van Devender, T.R. 1976. Foods of the desert tortoise, Gopherus agassizii, in Arizona and Utah. Herpetologica. 32(3): 247-251.
Hansen, R.M.; Martin, P.S. 1973. Ungulate diets in the Lower Grand Canyon. Journal of Range Management. 26(5): 380-381.
Hay F.R.; Probert, R.J. 2011. Chapter 20: Collecting and handling seeds in the field. In: Guarino, L.; Ramanatha, V.; Goldberg, E. Collecting plant genetic diversity: Technical Guidelines-2011 update. Rome, Italy: Bioversity International. 33 p.
Hill, A.C.; Hill, S.; Lamb, C.; Barrett, T.W. 1974. Sensitivity of native desert vegetation to SO2 and to NO2 combined. Journal of the Air Pollution Control Association. 24(2): 153-157.
Hodgkinson, H.S. 1983. Relationship between Cutler mormon-tea Ephedra cutleri and coppice dunes in determining range trend in northeastern Arizona. Journal of Range Management. 36(3): 375-377.
Holmgren, N.H.; Holmgren, P.K.; Cronquist, A. 2005. Intermountain flora: Vascular plants of the Intermountain West, U.S.A. Volume Two, Part B: Subclass Dilleniidae. Bronx, NY: New York Botanical Garden Press. 488 p.
Huffman, D.W.; Stoddard, M.T.; Springer, J.D.; Crouse, J.E.; Chancellor, W.W. 2013. Understory plant community responses to hazardous fuels reduction treatments in pinyon-juniper woodlands of Arizona, USA. Forest Ecology and Management. 289: 478-488.
Jameson, D.A.; Williams, J.A.; Wilton, E.W. 1962. Vegetation and soils of Fishtail Mesa, Arizona. Ecology. 43(3): 403-410.
Kearney, T.H. 1935. The North American species of Sphaeralcea subgenus Eusphaeralcea. Berkeley, CA: University of California Press. 127 p.
Kramer, A.; Foxx, A. 2016. Propagation protocol for production of propagules (seeds, cuttings, poles, etc.) Machaeranthera canescens (Pursh) A. Gray seeds. Native Plant Network. U.S. Department of Agriculture, Forest Service, National Center for Reforestation, Nurseries, and Genetic Resources. http://NativePlantNetwork.org [Accessed 2017 July 21].
Krausman, P.R.; Leopold, B.D.; Seegmiller, R.F.; Torres, S.G. 1989. Relationships between desert bighorn sheep and habitat in western Arizona. Wildlife Monographs. 102: 3-66.
La Duke, J. 2016. 48. Sphaeralcea. In: Flora of North America Editorial Committee, ed. Flora of North America North of Mexico. Volume 6 Malvaceae. http://www.efloras.org
Lady Bird Johnson Wildflower Center [LBJWC]. 2023. Sphaeralcea parvifolia A. Nelson. Native Plant Database. Austin, TX: Lady Bird Johnson Wildflower Center. https://www.wildflower.org/plants-main [Accessed 2023 June 23].
Lambert, S. 2005. Guidebook to the seeds of native and non-native grasses, forbs and shrubs of the Great Basin. Boise, ID: U.S. Department of the Interior, Bureau of Land Management, Idaho State Office. 136 p.
Love, S.L.; Akins, C.J. 2020. Sixth summary of the native seed germination studies of Norman C Deno: Species with names beginning with letters R through Z. Native Plants Journal. 21(2): 150-187.
Meyer, T.; Vernon, J. 2008. Native plant material development and seed and seeding technology for native Great Basin forbs and grasses. In: Shaw, N.; Pellant, M., eds. Great Basin Native Plant Selection and Increase Project: 2007 Progress Report. Boise, ID: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station: 7-24.
Moerman, D. 2003. Native American ethnobotany: A database of foods, drugs, dyes, and fibers of Native American peoples, derived from plants. Dearborn, MI: University of Michigan. http://naeb.brit.org
Monsen, S.B.; Stevens, R. 2004. Seedbed preparation and seeding practices. In: Monsen, S.B.; Stevens, R.; Shaw, N.L., comps. Restoring western ranges and wildlands, vol. 1. Gen. Tech. Rep. RMRS-GTR-136-vol-1. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station: 121-154.
Munz, P.A.; Keck, D.D. 1973. A California flora and supplement. Berkeley, CA: University of California Press. 1905 p.
Ogle, D.; Tilley, D.; Cane, J.; St. John, L.; Fullen, K.; Stannard, M.; Pavek, P. 2011. Plants for pollinators in the Intermountain West. Plant Materials Tech. Note 2A. Boise, ID: U.S. Department of Agriculture, Natural Resources Conservation Service. 40 p.
Ogle, D.; Tilley, D.; Cane, J.; St. John, L.; Fullen, K.; Stannard, M.; Pavek, P. 2019. Plants for pollinators in the Intermountain West. Plant Materials Tech. Note 2A. Boise, ID: U.S. Department of Agriculture, Natural Resources Conservation Service. 40 p.
Omernik, J.M. 1987. Ecoregions of the conterminous United States. Map (scale 1:7,500,000). Annals of the Association of American Geographers. 77(1): 118-125.
Owen, S.M.; Sieg, C.H.; Gehring, C.A. 2011. Rehabilitating downy brome (Bromus tectorum)-invaded shrublands using imazapic and seeding with native shrubs. Invasive Plant Science and Management. 4(2): 223-233.
Parkinson, H. 2003. Landscaping with native plants of the Intermountain Region. Tech. Ref. 1730-3. Boise, ID: U.S. Department of the Interior, Bureau of Land Management. 46 p.
Pendery, B.M.; Rumbaugh, M.D. 1986. Globemallows: Forbs for Utah rangelands. Utah Science: 41-45.
Pendery, B.M.; Rumbaugh, M.D. 1990. Survival and growth of globemallow (Sphaeralcea) species in dry land spaced-plant nurseries. Journal of Range Management. 43(5): 428-432.
Pendery, B.M.; Rumbaugh, M.D. 1993. Globemallows. Rangelands. 15(3): 127-130.
Plant Conservation Alliance [PCA]. 2015. National seed strategy for rehabilitation and restoration 2015-2020. Washington, DC: U.S. Department of the Interior, Bureau of Land Management. 52 p.
Richardson, B.; Kilkenny, F.; St. Clair, B.; Stevenson-Molnar, N. 2020. Climate Smart Restoration Tool. https://climaterestorationtool.org/csrt/
Rumbaugh, M.D.; Mayland, H.F.; Pendery, B.M.; Shewmaker, G.E. 1993a. Element concentrations in globemallow herbage. Journal of Range Management. 46(2): 114-117.
Rumbaugh, M.D.; Mayland, H.F.; Pendery, B.M.; Shewmaker, G.E. 1993b. Utilization of globemallow (Sphaeralcea) taxa by sheep. Journal of Range Management 46(2): 103-109.
Schmutz, E.M.; Michaels, C.C.; Judd, B.I. 1967. Boysag Point: A relict area on the North Rim of Grand Canyon in Arizona. Journal of Range Management. 20(6): 363-369.
Seglias, A.E.; Williams, E.; Bilge, A.; Kramer, A.T.; Peruzzi, L. 2018. Phylogeny and source climate impact seed dormancy and germination of restoration-relevant forb species. PLOS One. 13(2): 7-18.
SEINet – Regional Networks of North American Herbaria Steering Committee [SEINet]. 2017. SEINet Regional Networks of North American Herbaria. https://symbiota.org/seinet
Shock, C.; Feibert, E. 2016. Irrigation water requirements for native wildflower seed production. In: Kilkenny, F.; Edwards, F.; Malcomb, A., eds. Great Basin Native Plant Project: 2015 Progress Report. Boise, ID: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station: 175-176.
Shock, C.; Feibert, E.; Saunders, L.; Shaw, N. 2008. Subsurface drip irrigation for native wildflower seed production. In: Shock, C., ed. Malheur Experiment Station Annual Report 2007. Special Report 1087. Corvallis, OR: Oregon State University: 183-196.
Shock, C.; Feibert, E.; Saunders, L.; Shaw, N.; Sampangi, R.S. 2012. Irrigation requirements for native wildflower seed production. In: Shock, C., ed. Malheur Experiment Station Annual Report 2011. OSU AES Ext/CrS 141. Corvallis, OR: Oregon State University: 139-160.
Shock, C.C.; Feibert, E.B.G.; Rivera, A.; Saunders, L.D.; Shaw, N.; Kilkenny, F. 2015. Globemallow seed production is unresponsive to irrigation in a semi-arid environment. In: Shock, C.C., ed. Malheur Experiment Station Annual Report 2014. OSU AES Ext/ CrS152. Corvallis, OR: Oregon State University: 247-251.
Society for Ecological Restoration; International Network for Seed Based Restoration; Royal Botanic Gardens [SER, INSR, RBGK]. 2023. Seed Information Database (SID). Available from: https://ser-sid.org/
Sriladda, C.; Kratsch, H.A.; Larson, S.R.; Kjelgren, R.K. 2012. Morphological and genetic variation among four high desert Sphaeralcea species. Hortscience. 47(6): 715-720.
Tilley, D.; Taliga, C.; Burns, C.; St. John, L. 2013. Plant materials for pollinators and other beneficial insects in eastern Utah and western Colorado. Tech. Note 2C. Boise, ID: U.S. Department of Agriculture, Natural Resources Conservation Service. 54 p.
USDA Forest Service, Bend Seed Extractory [USDA FS BSE]. 2017. Nursery Management Information System Version 4.1.11. Local Source Report 34-Source Received. Bend, OR: U.S. Department of Agriculture, Forest Service, Bend Seed Extractory.
USDA Forest Service, Bend Seed Extractory [USDA FS BSE]. 2023. Nursery Management Information System Version 4.1.11. Local Source Report 34-Source Received. Bend, OR: U.S. Department of Agriculture, Forest Service Bend Seed Extractory.
USDA Forest Service, Great Basin Native Plant Project [USFS GBNPP]. 2014. Seed weight table calculations made in-house. Report on file. Boise, ID: U.S. Department of Agriculture, Forest Service, Boise Aquatic Sciences Laboratory. Available: https://www.fs.fed.us/rm/boise/research/shrub/Links/Seedweights.pdf
USDA Forest Service, Western Wildland Environmental Threat Assessment Center [USFS WWETAC]. 2024. TRM Seed Zone Applications. Prineville, OR: U.S. Department of Agriculture, Forest Service, Western Wildland Environmental Threat Assessment Center. https://research.fs.usda.gov/pnw/products/dataandtools/seed-zone-webmap
USDA Natural Resources Conservation Service [USDA NRCS]. 2025. The PLANTS Database. Greensboro, NC: U.S. Department of Agriculture, Natural Resources Conservation Service, National Plant Data Team. https://plants.sc.egov.usda.gov/
USDI Bureau of Land Management, Seeds of Success [USDI BLM SOS]. 2017. Seeds of Success collection data. Washington, DC: U.S. Department of the Interior, Bureau of Land Management, Plant Conservation and Restoration Program.
USDI Bureau of Land Management, Seeds of Success [USDI BLM SOS]. 2023. Bureau of Land Management technical protocol for the collection, study, and conservation of seeds from native plant species for Seeds of Success. Washington, DC: U.S. Department of the Interior, Bureau of Land Management, Plant Conservation and Restoration Program. 45 p.
USDI Bureau of Land Management, Seeds of Success [USDI BLM SOS]. 2024. Seeds of Success collection data. Washington, DC: U.S. Department of the Interior, Bureau of Land Management, Plant Conservation and Restoration Program.
USDI Environmental Protection Agency [USDI EPA]. 2017. Ecoregions. Washington, DC: U.S. Department of the Interior, Environmental Protection Agency. https://www.epa.gov/eco-research/ecoregions
Utah Crop Improvement Association [UCIA]. 2015. How to be a seed connoisseur. Logan, UT: UCIA, Utah Department of Agriculture and Food, Utah State University and Utah State Seed Laboratory. 16 p.
Vernon, J.; Meyer, T.; Walker, S.C. 2005. Native plant material development and seed and seeding technology for native Great Basin forbs and grasses. In: Shaw, N.L.; Pellant, M., eds. Great Basin Native Plant Project: 2004 Progress Report. Boise, ID: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station: 14-17.
Vernon, J.; Meyer, T.; Whittaker, A. 2006. Native plant material development and seed and seeding technology for native Great Basin forbs and grasses. In: Shaw, N.L.; Pellant, M., eds. Great Basin Native Plant Project: 2005 Progress Report. Boise, ID: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station: 20-24.
Weber, S.A.; Seaman, P.D., eds. 1985. Havasupai habitat: A.F. Whiting’s ethnography of a traditional Indian culture. Tucson, AZ: University of Arizona Press. 288 p.
Weber, W.A. 1976. Rocky Mountain flora. Boulder, CO: Colorado Associated University Press. 479 p.
Welsh, S.L.; Atwood, N.D.; Goodrich, S.; Higgins, L.C., eds. 2015. A Utah Flora. Fifth Edition, revised. Provo, UT: Brigham Young University. 990 p.
Young, J.A.; Palmquist, D.E. 1992. Plant age/size distributions in black sagebrush (Artemisia nova): Effects on community structure. The Great Basin Naturalist. 52(4): 313-320.
Young, S.; Bouck, M. 2007. Establishment and maintenance of the buy-back program for certified seed. In: Shaw, N.L.; Pellant, M., eds. Great Basin Native Plant Project: 2006 Progress Report. Boise, ID: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station: 120-126.
Young, S.A.; Schrumpf, B.; Amberson, E. 2003. The Association of Official Seed Certifying Agencies (AOSCA) native plant connection. Moline, IL: AOSCA. 9 p. Available: https://seedcert.oregonstate.edu/sites/seedcert.oregonstate.edu/files/pdfs/aoscanativeplantbrochure.pdf
How to Cite
Gucker, Corey L.; Shaw, Nancy L. 2025. Small-leaf globemallow (Sphaeralcea parvifolia). In: Gucker, C.L.; Shaw, N.L., eds. Western forbs: Biology, ecology, and use in restoration. Reno, NV: Great Basin Fire Science Exchange. Online: https://westernforbs.org/species/small-leaf-globemallow-sphaeralcea-parvifolia/