Authorship
Gucker, Corey L.; Shaw, Nancy L.
Publication Date
September 2024
Nomenclature
Spreading fleabane (Erigeron divergens [Torrey & A. Gray]) belongs to the Astereae tribe within the Asteraceae family (Nesom 2006).
Family
Asteraceae – Aster family
Genus
Erigeron
Species
divergens
NRCS Plant Code
ERDI4 (USDA NRCS 2024).
Subtaxa
None (Nesom 2006).
Synonyms
Erigeron accedens Greene, E. incomptus A. Gray, E. solisaltator G. L. Nesom (Nesom 2006).
Common Names
Spreading fleabane, branching fleabane, fleabane daisy, spreading daisy (Welsh et al. 2016; LBJWC 2022; USDA NRCS 2024).
Chromosome Number
2n = 18, 27, 36 (Pinkava and Keil 1977; Strother 1983; Nesom 2015). 2n = 54 was also reported (Löve 1974). Polyploidy is common for the species. Diploids are scattered through at least the southern part of the species range (Nesom 2006).
Hybridization
Hybrids occur between spreading fleabane and running fleabane (E. tracyi), trailing fleabane (E. flagellaris), plains fleabane (E. modestus), and possibly other close relatives (Nesom 2015).
Distribution
Spreading fleabane is widely distributed throughout the western United States and northwestern Mexico (Nesom 2015) and is most widespread in the interior West (Cronquist et al. 1994). Populations occur in British Columbia and Alberta south through all states west of North Dakota and Texas. In Mexico, populations occur in Baja California, Chihuahua, Coahuila, Durango, Sonora, and likely in the more southern states (Nesom 2006). In the United States, distribution is limited at the eastern edge of its range with populations occurring only in the western parts of the Dakotas, Nebraska, Kansas, Oklahoma, and far west Texas (USDA NRCS 2024). Diploid forms of the species are scattered through at least the southern part of the species’ range (Nesom 2015).
Habitat And Plant Associations
Spreading fleabane grows in a variety of dry to moist habitats including riverbanks, meadows, desert scrublands, desert to semi-arid grasslands, woodlands, forests, and disturbed sites (Figs. 1, 2). Common associates include saltbush (Atriplex spp.), blackbrush (Coleogyne ramosissima), rabbitbrush (Chrysothamnus spp.), sagebrush (Artemisia spp.), pinyon-juniper (Pinus–Juniperus spp.), oak (Quercus spp.), ponderosa pine (Pinus ponderosa), quaking aspen (Populus tremuloides), and spruce (Picea spp.) (Hickman 1993; Cronquist et al. 1994; Nesom 2006; Spellenberg 2012; Riser and Meyers 2020). In the Canyonlands and Colorado Plateau of Utah, Colorado, and Arizona, spreading fleabane is common in lowland riparian, cool desert shrub, and pinyon-juniper communities (Fagan 1998).

Figure 1. Spreading fleabane growing with cheatgrass (Bromus tectorum) and prickly lettuce (Lactuca serriola) in a weedy area in California. Photo: USDI, Bureau of Land Management (BLM) CA320, Seeds of Success (SOS).
Following are descriptions of forest, shrubland, grassland, and riparian vegetation communities where spreading fleabane grows or once grew. These communities are scattered throughout the range of the species but their mention here does not necessarily indicate species preference or regularity over other communities not mentioned.
Forests, woodlands, shrublands. In the Lake Tahoe region of California and Nevada, spreading fleabane was common in the understory of white fir (Abies concolor), Jeffrey pine (Pinus jeffreyi), and ponderosa pine forests just below subalpine vegetation (Smiley 1915). It also occurred in chaparral shrublands in California and Gambel oak (Quercus gambelii) thickets in Utah. In the Santa Ynez Mountains near Santa Barbara, California, spreading fleabane grew in openings or recently burned chamise (Adenostoma fasciculatum)-dominated chaparral (Christensen and Muller 1975). In Utah, it occurred from desert shrublands and valley grasslands into Gambel oak shrublands in the lower montane forest in the Wasatch Mountains (Hayward 1948).
Grasslands. Spreading fleabane was reported in xeric grasslands in Arizona and Colorado and in moist meadows in Colorado. At the Fort Huachuca Military Reservation in the Huachuca Mountains of Arizona, spreading fleabane was sampled in xeric grasslands in a wet year and in a very dry year. Total annual precipitation averaged 13 in (340 mm) in the wet year (2008) and 6.5 in (165 mm) in the dry year (2009). Precipitation in 2009 was the second lowest recorded in 60 years (Lance et al. 2017). In Colorado, spreading fleabane was considered characteristic of xerophytic grasslands (Ramaley 1919) and was abundant in a mixed-grass meadow near Boulder (Concilio et al. 2016). In western Oklahoma, spreading fleabane occurred between mats of grasses in needlegrass-grama (Achnatherum–Bouteloua spp.) mixed prairie and buffalograss (Bouteloua dactyloides)-grama shortgrass plains on moderately moist soils (Bruner 1931).

Figure 2. Spreading fleabane growing in a Montana grassland. Photo: USDI, BLM MT050, SOS.
Riparian. In Jackson Hole Wildlife Park, Wyoming, spreading fleabane was common in big sagebrush communities and streamside woodlands dominated by cottonwood (Populus spp.) (Reed 1952). It was also reported in surveys of the intermittent Hassayamp River in west-central Arizona (Drezner and Fall 2002).
Elevation
The elevation range for spreading fleabane is 160 to 9,500 ft (50-2,900 m) (Cronquist et al. 1994; Nesom 2006; Welsh et al. 2016; Riser and Meyers 2020). Reported ranges are 160 to 7,220 ft (50-2,200 m) for Oregon (Riser and Meyers 2020), 200 to 8,530 ft (60-2,600 m) for California (Munz and Keck 1973; Hickman 1993), 3,200 to 9,500 ft (975-2,900 m) for Utah (Welsh et al. 2016), and 200 to 8,900 ft (60-2,700 m) in the Sonoran Desert (Spellenberg 2012).
Soils
Spreading fleabane commonly occurs and can form dense populations on sandy or gravelly soils (Bowers 1980; Taylor 1992; Spellenberg 2001; Nesom 2006) in dry and waste places (Cronquist 1947). In a tamarisk/arrowweed (Tamarix ramosissima/Pluchea sericea) community along the Colorado River in the Grand Canyon between Lees Ferry and Diamond Creek, spreading fleabane grew on sandy soils with average daily inundation frequency of 0.16 (Stevens et al. 1995).
The following paragraph provides specifics related to soils where spreading fleabane grows and gives an approximation of tolerance but not necessarily preference. In the Santa Ynez Mountains near Santa Barbara, California, spreading fleabane occurred in recently burned chaparral or chaparral openings with shallow clay loam soils low in phosphorous and nitrogen (Christensen and Muller 1975). At Grants Lava Bed in west-central New Mexico, spreading fleabane grew in ponderosa pine forests on lava flows and loose volcanic deposits on gentle slopes (Lindsey 1951). In a study that evaluated topography, soils, and vegetation relationships in ponderosa pine forests in northern Arizona, spreading fleabane was most abundant in clay basalt/snakeweed (Gutierrezia spp.) and heavily grazed park/aster (Symphyotrichum spp.) ecosystems. Soils encountered included black cinders, red cinders, clay basalt, mesic basalt, mesic limestone, rocky basalt, xeric basalt, and xeric limestone, and spreading fleabane grew in all but the cinder soils (Abella and Covington 2006). Spreading fleabane was abundant in a mixed-grass meadow dominated by western wheatgrass (Pascopyrum smithii), buffalograss, grama, and some cheatgrass (Bromus tectorum) near Boulder, Colorado. This vegetation occupied well drained colluvial sandy loams at sites with an average annual precipitation range of 11 to 34 in (277-867 mm) and an average temperature of 51.6 °F (10.9 °C) (Concilio et al. 2016).
Description
Spreading fleabane is a polymorphic and morphologically variable species due to mixed diploid and polyploid populations, hybridization, and asexual reproduction (Nesom 2015). The species grows as an annual, biennial, or perennial. In the southwestern part of its range there are two annual types, one that is mostly single stemmed from the base and one with multiple stems from the base. Short-lived perennials are also common in the southern part of the species range. More northern populations are primarily annual. Perennials flower in the first year from a slender taproot but later the taproot becomes woody as does the caudex (Fig. 3) (Nesom 2015). In the Chihuahuan Desert where annual precipitation is bimodal, spreading fleabane is considered a winter annual (Guo et al. 2002).

Figure 3. Spreading fleabane pulled from a grassland in Montana. This plant appears to be a perennial, although this lifeform is thought to be less common in northern parts of the species range. Photo: USDI, BLM MT050, SOS.
Plants develop single to multiple, erect, ascending to decumbent stems 2.8 to 28 in (7-70 cm tall) (Fig. 4) (Cronquist 1947; Nesom 2006). Stems are branched at or near the base and then freely upwards (Cronquist 1947; Welsh et al. 2016). Plants differ in hairiness from densely pilose to sparsely hirsute with straight to curled hairs that are spreading, descending, or ascending (Cronquist 1947; Munz and Keck 1973; Hickman 1993; Nesom 2006; Spellenberg 2012; Hitchcock and Cronquist 2018; Riser and Meyers 2020; LBJWC 2022).
Leaves are alternate, hirsute, basal and cauline (Cronquist et al. 1994; Nesom 2006). Basal leaves are obovate-spatulate, petiolate, 0.4 to 2.8 in (1-7 cm) long, and 4 to 14 mm wide. Their margins are entire or with two to three pairs of teeth or lobes (Munz and Keck 1973; Cronquist et al. 1994; Nesom 2006; Spellenberg 2012; Riser and Meyers 2020). Basal leaves are often deciduous and lost by the time of flowering (Hitchcock and Cronquist 2018). Stem leaves are numerous, smaller, oblong to linear, and mostly less than 2.4 in (6 cm) long and 5 mm wide, decreasing in size and becoming sessile upwards (Munz and Keck 1973; Hickman 1993; Cronquist et al. 1994; Lesica 2012; Welsh et al. 2016; Hitchcock and Cronquist 2018).

Figure 4. Spreading fleabane growing on the Colorado Plateau. Photo: USDI, BLM CP1, SOS.
Flower heads are several to numerous (20+), diffuse, flat-topped, corymb-like arrays (Hickman 1993; Cronquist et al. 1994; Hitchcock and Cronquist 2018; Riser and Meyers 2020). Each includes 75 to 150 pistillate ray florets surrounding hundreds of bisexual disk florets (Fig. 5) (Hickman 1993; Hitchcock and Cronquist 2018; Riser and Meyers 2020). Ray florets are white to lavender to blue, 5 to 10 mm long, 0.5 to 1.2 mm wide, and sometimes scarcely developed or lacking (Hickman 1993; Spellenberg 2001; Welsh et al. 2016; Hitchcock and Cronquist 2018). Disk florets are yellow and 2 to 3 mm long (Hitchcock and Cronquist 2018). Involucres (4-5 mm tall) include two to four whorls of non-overlapping bracts (Munz and Keck 1973; Cronquist et al. 1994; Spellenberg 2001; Nesom 2006; Riser and Meyers 2020; LBJWC 2022). Bracts are mostly side by side and not overlapping (Spellenberg 2001).

Figure 5. Spreading fleabane flower heads. Photo: USDI, BLM MT050, SOS.
Spreading fleabane fruits are flattened achenes 0.9 to 1.2 mm long (Cronquist et al. 1994; Nesom 2006). Achenes have sparsely strigose faces and two to four whitish nerves (Cronquist et al. 1994; Riser and Meyers 2020). Achenes, hereafter referred to as seeds, have double pappi with 5 to 12 fragile inner barbellate bristles surrounded by minute bristles or setose scales (Hickman 1993; Cronquist et al. 1994; Spellenberg 2012; Welsh et al. 2016; Riser and Meyers 2020).
Belowground Relationships And Interactions
Trappe et al. (1981) reported the presence of mycorrhizae in root systems of spreading fleabane in semi-arid or arid environments.
Reproduction
Spreading fleabane occurs as sexual and asexually reproducing plants (Keil and Pinkava 1976). Asexual plants produce seed apomictically from diploid cells without fertilization. These develop into clones of the parent plant.
Phenology
Across the range of spreading fleabane, flowering can occur from February to November but is most common from April to October (Munz and Keck 1973; Nesom 2006; Spellenberg 2012). The flowering period is shorter in the northern part of the species’ range. In Oregon, flowering occurs from May to September (Riser and Meyers 2020), but in the Sonoran Desert it occurs from February to October (Bowers 1980; Spellenberg 2012).
Later flowering was observed with decreased precipitation over 20 years of observations along an elevation gradient from desert scrub (3,100 ft [945 m]) to oak woodlands (5,482 ft [1,671 m]) in the Santa Catalina Mountains of Tucson, Arizona. Spreading fleabane showed a significant trend of later onset of flowering date of about 5 days/year later from between 1984 and 2003. This trend coincides with decreased fall precipitation over the same time period that was likely an explanatory variable in delayed onset of flowering (Crimmins et al. 2010).
Pollination
Spreading fleabane flowers are visited and pollinated by a variety of insect and bee species. Flowers were visited by 11 pollinator taxa based on surveys of nine sites in western Montana that supported low and high densities of invasion by spotted knapweed (Centaurea stoebe). Sites were visited once a week from July 8 to August 21 for two 15-minute sessions between 9 am and 5 pm (Herron-Sweet et al. 2016). In a study that surveyed bees and flowers in hedgerows, agricultural fields, woodlots on farms, and unmanaged native woodland farms in southeastern Arizona, spreading fleabane was visited by 19% of the total native bee foragers in hedgerows in the summer monsoon sampling period (Hannon and Sisk 2009).
Ecology
Spreading fleabane is a ruderal species (Korb et al. 2004) often associated with disturbed sites (Spellenberg 2012; Riser and Meyers 2020). Abundance of the species is often greater on disturbed than undisturbed sites (Christensen and Muller 1975; Crawford et al. 2001; Metlen et al. 2006).
Seed And Seedling Ecology
Seedbank studies suggest that spreading fleabane is a prolific seed producer and its seed likely persists in the soil. A study of granivory in the Chihuahuan Desert suggests that more spreading fleabane seed is taken by ants than rodents (Samson et al. 1992).
Spreading fleabane was one of the most dominant ruderal species to emerge from soil samples collected 10 ft (3 m) outside of pile burn scars (Korb et al. 2004) created by thinning and slash burning in ponderosa pine forests near Flagstaff, Arizona. Slash piles were 30 ft (9 m) in diameter, 10 to 13 ft (3-4 m) tall, and burned in February 2000. Soil was collected in May 2000 and kept in an unheated greenhouse where seedling emergence was monitored (Korb et al. 2004).
Spreading fleabane seedlings also emerged from soil collected prior to and after tree thinning in northern Arizona ponderosa pine forests (Abella 2009b). Emergence from soil samples was monitored in a greenhouse where aqueous smoke solution treatments were also evaluated. Spreading fleabane emerged from all soil samples and treatments with no significant differences between any treatments or sites. Soil was collected in June or July, stored at 23 °F (-5 °C) for 5 to 6 months before evaluating emergence and testing aqueous smoke treatments (Abella 2009b).
Two seedbank studies reported emergence of spreading fleabane from soil depths of 2 to 4 in (5-10 cm), suggesting persistence in the soil (Springer 1999; Abella et al. 2007). Frequency of spreading fleabane seedlings was 26% and seedling density was 232/ft2 (2,500/m²) from 0 to 2-in (5 cm) soil cores (Abella et al. 2007). Seedling frequency was 17% and density was 77/ft2 (833/m2) from 2- to 4-in (5-10 cm) soil cores. Soil samples were collected from ponderosa pine forests near Flagstaff, Arizona, from mid-May to August. Spreading fleabane frequency was 94% in aboveground vegetation. The composition of the seedbank was determined using the emergence method. Emergence from the 0 to 2-in (0-5 cm) soil depths likely included current year seed production, but emergence from deeper soils suggested persistence of prior years’ seed production (Abella et al. 2007).
Spreading fleabane also emerged at low levels from mineral soil samples collected from 2 to 4 in (5-10 cm) deep in the Arizona strip between Utah and the Colorado River (Springer 1999). Soil samples were collected in mid-September in 1997 from ponderosa pine, Gambel oak, New Mexico locust (Robinia neomexicana), and big sagebrush (Artemisia tridentata) vegetation types. Soil was stored at 39 °F (4 °C) for 6 weeks before being put in a greenhouse where seedling emergence was monitored. Density of emerging spreading fleabane seedlings was 82/ft2 (883/m2) from big sagebrush, 2/ft2 (23/m2) from New Mexico locust and Gambel oak, and none from ponderosa pine soil samples. Following thinning treatments in the ponderosa pine forests, spreading fleabane emerged at low levels from mineral soil samples collected at 2 to 4 in (5-10 cm) deep, suggesting persistence in the seed bank (Springer 1999).
Granivory. In a study conducted in the Chihuahuan Desert of southeastern Arizona, density of spreading fleabane increased significantly (P < 0.01) on plots where desert harvester ants (Pogonomyrmex rugosus [or P. barbatus]) were removed but not on plots where kangaroo rats (Dipodomys spp.) were removed (Samson et al. 1992). The study monitored plant responses to treatments that removed seed-eating rodents and ants over a ten-year period (1978-1987). Treatments included: 1) removal of all granivorous ants with repeated insecticide treatments, 2) elimination of all seed-eating rodents by trapping and removal, 3) removal of both granivorous ants and rodents, 4) elimination of desert harvester ants by poisoning at colony entrances, 5) removal of all by trapping and removal (Samson et al. 1992).
Disturbance Ecology
Studies show that spreading fleabane is typically more abundant in openings than in closed canopy woodlands or forests and in recently disturbed or heavily grazed than in undisturbed vegetation (Gardner and Hubbell 1934; Christensen and Muller 1975; McArthur et al. 1988; Metlen et al. 2006; Abella 2009a).
In an area with active and inactive mounds (abandoned 4-15 yrs previously) created by banner-tailed kangaroo rats (D. spectabilis), spreading fleabane was one of the most conspicuous species occurring at the edge of inactive mounds, where it was more abundant than on inactive mounds (P < 0.01). The study area was the Chihuahuan Desert near Portal, Arizona, in transitional vegetation between shrublands and grasslands. Shrublands were dominated by whitethorn acacia (Acacia constricta), American tarwort (Flourensia cernua), longleaf jointfir (Ephedra trifurca), and broom snakeweed (Gutierrezia sarothrae), and grasslands were dominated by lovegrasses (Eragrostis spp.) and threeawns (Aristida spp.) (Guo 1996).
Shading. Spreading fleabane was much more frequent in openings than beneath increasing canopy cover in ponderosa pine-Gambel oak forests in northern Arizona (Abella 2009a). Understory plant communities and soil properties were compared for five canopy types with increasing canopy cover: openings (free of canopy cover), single ponderosa pine, single stem Gambel oak, dispersed clumps of Gambel oak, and Gambel oak thickets. Spreading fleabane occurred in all but the Gambel oak thickets, and frequency decreased with increasing canopy cover. Frequency was 64% in openings, 24% in single ponderosa pine, 14% in single stem Gambel oak, 8% in dispersed Gambel oak clumps, and 0% in Gambel oak thickets. Soil moisture and litter weight were significantly lower in openings than in any other canopy type (P < 0.01). Percent sand, silt, and clay did not differ significantly (Abella 2009a).
Fire. In most studies comparing the abundance of spreading fleabane abundance, there was more spreading fleabane on burned than unburned plots.
Cover of spreading fleabane increased significantly (P < 0.05) following thinning and burning treatments in ponderosa pine-Douglas-fir (Pseudotsuga menziesii) forests at the Lubrecht Experimental Forest near Missoula, Montana. Cover was not significantly different between pre-treatment levels, thinning alone, or burning alone treatments. The prescribed fires occurred in early May and were of low to moderate severity (Metlen et al. 2006). Spreading fleabane occurred in recently burned (1-2 yrs) but not in long unburned chamise-dominated chaparral in the Santa Ynez Mountains near Santa Barbara, California. The fire occurred on September 17, 1971, on moderately steep slopes (10-30%) with primarily southwestern exposures. Spreading fleabane was most common in heavily disturbed sites, which included a dirt road winding through the burned area (Christensen and Muller 1975). In ponderosa pine forests in central and northern Arizona, spreading fleabane cover was much greater on high-severity burned (2.9%) than unburned (0.1%) plots. This study evaluated the effects 2 years after three wildfires that burned in June and July on the Kaibab and Coconino National Forests (Crawford et al. 2001).
Spreading fleabane was an indicator species on severely burned plots 2 and 3 years after fire in ponderosa pine forests on the Apache Sitgreaves National Forest in Arizona (Shive et al. 2013). In a portion of the study area, thinning and pile burning occurred 12 years prior to the 2002 fire. Postfire effects were compared in thinned and untreated areas 2, 3, and 9 years after the fire. Spreading fleabane was an indicator (P < 0.05) of untreated burned plots where fire severity was most severe. The entire burned area was seeded in 2002 and 2003 with a mix of native and nonnative species that did not include spreading fleabane (Shive et al. 2013).
At the Fort Valley Experimental Forest in the Coconino National Forest, spreading fleabane was an indicator species (P < 0.05) of medium-intensity thinned then prescribe burned sites (Stoddard et al. 2011). Indicator species were those most abundant and frequent in a particular treatment. Thinning occurred in 1999, and the prescribed fire occurred in 2000. Thinning treatments were high intensity, medium-intensity, low-intensity, and no thinning. Spreading fleabane was an indicator of medium-intensity thinned and burned forests in 2002 (Stoddard et al. 2011). In the Apache Sitgreaves National Forest in eastern Arizona, spreading fleabane was an indicator species of post-fire but not pre-fire warm-dry mixed conifer forests. The fire occurred in early June 2011. Spreading fleabane was a significant indicator (P < 0.05) 1 and 5 years after fire (Springer et al. 2018).
Short-term density reductions of spreading fleabane followed burning at the Appleton-Whittell Research Ranch in southeastern Arizona (Table 1, Bock and Bock 1992). The study occurred in plains lovegrass (E. intermedia) grassland with mimosa (Mimosa spp.) shrubs. Plots were measured before and for 2 years after a mid-June prescribed fire in 1984 (Bock and Bock 1992).
Table 1. Density (stems/1 ft2) of spreading fleabane after a prescribed fire (June 12, 1984) in southeastern Arizona (Bock and Bock 1992).
| Year | Burned | Unburned |
| Density of spreading fleabane (stems/ft2) | ||
| 1983 (prefire) | 0.7a | 0.96a |
| 1984 (postfire yr 1) | 0.05b | 0.67a |
| 1985 (postfire yr 2) | 0.75a | 0.44a |
Values within the same row followed by different letters were significantly different (P ≤ 0.001).
At 10 years postfire, spreading fleabane was an indicator of control plots (P = 0.017) in a study that evaluated thinning and burning and burning treatments in mixed-conifer forests in the San Juan Mountains of southwestern Colorado (Korb et al. 2020). At that time, canopy fuels were significantly lower in thinned and burned than in control and burned plots. Forests prior to treatment included ponderosa pine, Douglas-fir, and white fir and occurred on moderately steep (15-30%) southern slopes. Thinning occurred in 2004 and thinned and unthinned plots were prescribed burned in 2007 or 2008 (Korb et al. 2020).
Grazing. Studies reported high frequencies of spreading fleabane in heavily grazed mountain big sagebrush and ponderosa pine communities (Gardner and Hubbell 1934; McArthur et al. 1988). Frequency of spreading fleabane was 73% in heavily mule deer (Odocoileus hemionus) browsed and 58% in unbrowsed mountain big sagebrush (Artemisia tridentata subsp. vaseyana) vegetation just south of the Utah-Juab County line in Utah. The browsed and largely unbrowsed plots were bisected by a deer fence (McArthur et al. 1988). In overgrazed vegetation on nearly level openings in ponderosa pine forests, frequency of spreading fleabane on heavily grazed plots was more than double that on plots protected for 8 years (Gardner and Hubbell 1934). Frequency was also greater on lightly grazed than protected plots. The study occurred at the Navajo Experiment Station in northwestern New Mexico after 8 years of protection from or reductions in grazing intensity. Grazing was primarily by sheep (~85%) and the rest by horses and cattle prior to and after 1934 (Gardner and Hubbell 1934).
Wildlife And Livestock Use
Spreading fleabane can be a food source for large ungulates, domestic livestock, rodents, game birds, and insects.
Large mammals. Through a literature review, Krausman et al. (1978) found that spreading fleabane comprised a low proportion (up to 5%) of winter mule deer diets. In hills near Springville, Utah, spreading fleabane occurred in the diets of both elk (Cervis canadensis) and mule deer (Mower and Smith 1989). The study analyzed diet composition from fecal samples collected from January to March in 1983, where spreading fleabane made up small percentages of mule deer and elk diets (Mower and Smith 1989). Spreading fleabane occurred in 21% of samples at a high volume of 3% in late winter (26 February-24 March) diets of white-tailed deer (O. virginianus) (McCulloch 1973). The study analyzed stomachs of 14 white-tailed deer over a period of 5 years from the Three Bar Wildlife area of the Tonto National Forest, Arizona, in a mix of chaparral and desert vegetation (McCulloch 1973). Both mule deer and white-tailed deer fed on spreading fleabane in Big Bend National Park, Texas (Krausman 1978). Stomach contents were analyzed seasonally between June 1972 and April 1974 for 25 white-tailed and 29 mule deer. The volume of spreading fleabane was a maximum of 5.4% in white-tailed deer diets and 2.4% in mule deer diets (Krausman 1978).
Spreading fleabane is important to pronghorn (Antilocapra americana) (Buechner 1950; Schwartz and Nagy 1976). Based on an analysis of fecal samples, spreading fleabane was the second most common plant in June pronghorn diets in the Central Plains Experimental Range in northeastern Colorado. Samples were taken from blue grama (Bouteloua gracilis)-buffalograss short-grass prairie in March, May, June, August, and October. Spreading fleabane was only recovered from fecal samples collected in June (Schwartz and Nagy 1976). In the Trans-Pecos region of Texas, spreading fleabane is considered good forage and utilized by pronghorn (Buechner 1950).
Spreading fleabane is considered more palatable to sheep and goats than to cattle or horses (U.S. Department of Agriculture 1937; Humphrey 1955). On the Central Plains Experimental Range north of Greely, Colorado, cattle diets were determined by esophageal fistula and fecal sample analyses. Collections were made for 2 years in June, July, and August on lightly, moderately, and heavily grazed sites. Spreading fleabane was not found in diets in 1969 but in 1971 it comprised 4% of diets by weight of esophageal fistula samples and 1% of fecal samples (Varva et al. 1978).
Small mammals. A variety of small mammals feed on spreading fleabane. Based on monthly sampling of shortgrass prairie for one year in northeastern Colorado, spreading fleabane made up a trace of the dry weight northern grasshopper mice (Onychomys leucogaster) and thirteen-lined ground squirrels (Spermophilus tridecemlineatus) diets (Flake 1973). Spreading fleabane was also identified in the diets of white-tailed and black-tailed jackrabbits (Lepus townsendii and L. californicus) diets in the same area, where it made up a high of 2% dry weight in white-tailed jackrabbit April diets (Flinders and Hansen 1972).
In ponderosa pine forests in the Coconino National Forest of Arizona, spreading fleabane was important in the diets of small rodents. The study was conducted in the spring and summer over 3 years and the small rodents included deer mice (Peromyscus maniculatus), brush mice (P. boylei), Mexican woodrats (Neotoma mexicana), southern pocket gophers (Thomomys umbrinus), chipmunks (Eutamias spp.), and golden-mantled ground squirrels (Spermophilus lateralis) (Goodwin and Hungerford 1979).
Birds and insects. Spreading fleabane is important in greater sage-grouse (Centrocercus urophasianus) habitats in the Great Basin (Lambert 2005) and utilized as a host or for feeding by a variety of insects and pollinators.
Spreading fleabane is a food source for redlegged grasshoppers (Melanoplus femurrubrum) (Capinera 2001, 2020). Adult metallic wood-boring beetles (Acmaeodera quadrivittatoides) feed on spreading fleabane flowers (Nelson and Westcott 1995). It is a potential nectar host plant for Mount Charleston blue butterfly (Icaricia [Plebejus] shasta charlestonensis) (Carvajal-Acosta et al. 2015), and utilized for feeding or as a host by northern checkerspot (Chlosyne palla) and field crescent (Phyciodes pulchella) butterflies (James and Nunnallee 2011). It is a host plant for Cucullia lilacina moths (Robinson et al. 2024), and visited by many bee species (Asmeadiella bucconis, A. cactorum, A. melioloti, Calliopsis fracta, Chelostoma minutum, Dianthidium singular, D. ulkei, Heriades cressoni, Megachile brevis, M. fidelis, and Perdita rivalis) (Discover Life 2024).
In a study that surveyed bee and flower relationships in southeastern Arizona, spreading fleabane was visited by 19% of total native bee foragers in hedgerows sampled during the summer monsoon (Hannon and Sisk 2009). A large micro bee fly species (Mythicomya sedonae) endemic to the Sedona area of Arizona, was first discovered in a collection from spreading fleabane. The bee fly was not observed until 1:30 pm, though other Bombyliidae visited and fed on spreading fleabane flowers earlier in the day (Evenhuis 1989).
Ethnobotany
Erigeron species and spreading fleabane specifically have ceremonial and medical uses for western Indigenous tribes. Thompson Indians of British Columbia chewed Erigeron leaves for sore throats and made a salve from toasted or fresh leaves and grease. The salve was used for pains, sores, swellings, and wounds (Turner et al. 1990). Other Canadian First Nations people made a tea using whole Erigeron plants to treat fevers, bronchitis, hemorrhoids, and cough. They used the plants as a diuretic to treat kidney stones, painful urination, diarrhea, gout, and diabetes. They ground dried Erigeron flowers into a powder and snorted it for fevers and headaches or inhaled it for head colds (MacKinnon et al. 2014).
Kiowa Indians of the Plains and Black Hills considered spreading fleabane an omen of good fortune when brought into the home (Vestal and Schultes 1939). The Navajo added spreading fleabane to warm water and gave it to pregnant women to aid delivery (Elmore 1944). Kayenta Navajo used spreading fleabane as a snuff for headaches (Wyman and Harris 1951). Ramah Navajo used it ceremonially and considered roots of spreading fleabane a life medicine. They also mixed spreading fleabane with other plants to make a cold infusion eyewash or to treat snake bites (Vestal 1952). In Mexico, spreading fleabane was used to treat ulcers and internal injuries (Gonzalez et al. 2004 cited in Rodríguez-Chávez et al. 2017).
Horticulture
Spreading fleabane seeds are available commercially. It is drought tolerant, grows in a variety of soils, attracts pollinators, and produces showy flowers. Mixing the very tiny seeds with sand was a method recommended for broadcasting the seed (Grand Prismatic Seed 2024).
Revegetation Use
Although use of spreading fleabane for revegetation or restoration was not reported in the available literature (2024), its ruderal habit, drought tolerance, and growth on many soils suggest it could be an important restoration species in the future (Korb et al. 2004; Abella and Covington 2006; Lance et al. 2017). In northwestern New Mexico, spreading fleabane grew on untreated coal mine spoils about 3 miles (5 km) east of Windowrock, Arizona. It was not common but established as a volunteer by 1974 on sites that were mined between 1963 and 1966 (Wagner et al. 1978).
Developing A Seed Supply
For restoration to be successful, the right seed needs to be planted in the right place at the right time. Coordinated planning and cooperation is required among all partners to select appropriate species and seed sources; these are determined by site conditions and current or projected revegetation requirements and goals (PCA 2015) and consultation with land managers and stakeholders.
Developing a seed supply begins with seed collection from native stands. Collected seed is distributed for restoration or production of nursery stock. Increase in agricultural seed fields may be required to provide a supply of seed (PCA 2015). Production of nursery stock requires less seed than large-scale seeding operations. Regardless of the size and complexity of any revegetation effort, seed certification is essential for tracking seed origin from collection through use (UCIA 2015).
Seed Sourcing
Because empirical seed zones are not currently available for spreading fleabane, generalized provisional seed zones developed by Bower et al. (2014) may be used to select and deploy seed sources. These provisional seed zones identify areas of climatic similarity with comparable winter minimum temperature and aridity (annual heat to moisture index). In Figure 6, Omernik Level III Ecoregions (Omernik 1987) overlay the provisional seed zones to identify climatically similar but ecologically different areas. For site-specific disturbance regimes and restoration objectives, seed collection locations within a seed zone and ecoregion may be further limited by elevation, soil type, or other factors.
The Western Wildland Environmental Threat Assessment Center’s (USDA FS WWETAC 2017) Threat and Resource Mapping (TRM) Seed Zone application provides links to interactive mapping features useful for seed collection and deployment planning. The Climate Smart Restoration Tool (Richardson et al. 2020) and the Southwest Deserts Provisional Seed Transfer Zones (Shryock et al. 2021) can also guide revegetation planning, seed collection, and seed deployment, particularly when addressing climate change considerations.
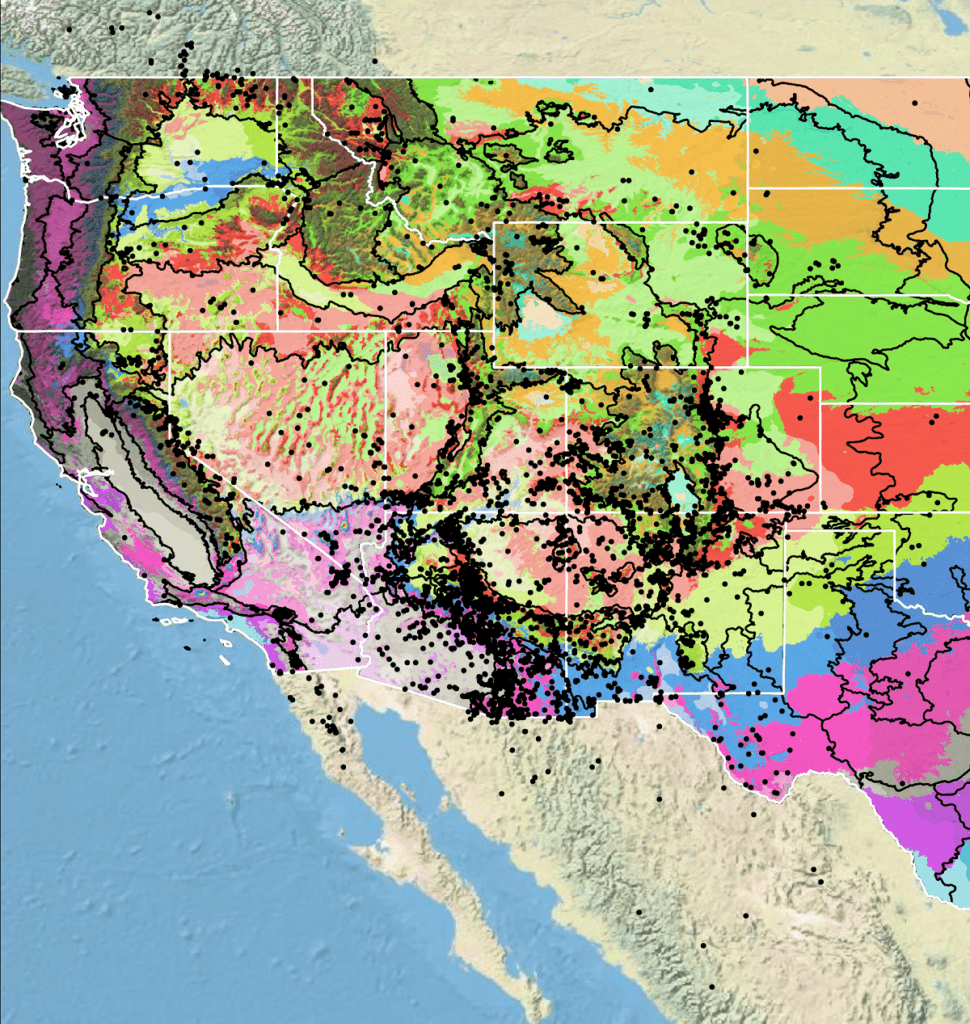
Figure 6. Distribution of spreading fleabane (black circles) based on geo-referenced herbarium specimens and observational data from 1871-2023 (CPNWH 2017; SEINet 2017; USDI USGS 2020). Generalized provisional seed zones (colored regions) (Bower et al. 2014) are overlain by Omernik Level III Ecoregions (black outlines) (Omernik 1987; USDI EPA 2018). Interactive maps, legends, and a mobile app are available (USDA FS WWETAC 2017; https://www.fs.usda.gov/wwetac/Seedzonemapping.php). Map prepared by Sarah Barga, USFS RMRS.
Releases
As of 2024, there were no spreading fleabane germplasm releases.
Wildland Seed Collection
Spreading fleabane seed is wind dispersed, and indeterminate seed production occurs over a broad window of time. Population monitoring is important for timing the collection of mature seed (Fig. 7).

Figure 7. Spreading fleabane seedhead and seeds with pappus (scale is cm). Photo: USDI, BLM MT050, SOS.
Wildland Seed Certification
Verification of species and tracking of geographic source is necessary whether wildland seed is collected for immediate project use or as stock seed for cultivated increase. This official Source Identification process can be accomplished by following procedures established by the Association of Official Seed Certifying Agencies (AOSCA) Pre-Variety Germplasm Program (UCIA 2015; Young et al. 2020). Wildland seed collectors should become acquainted with state certification agency procedures, regulations, and deadlines in the states where they collect.
If wildland-collected seed is to be sold for direct use in ecological restoration projects, collectors must apply for Source-Identified certification prior to making collections. Pre-collection applications, site inspections, and species and seed amount verification are handled by the AOSCA member state agency where seed collections will be made (see listings at AOSCA.org).
If wildland seed collected by a grower or private collector is to be used as stock seed for planting cultivated seed fields or for nursery propagation (See Agricultural Seed Field Certification section), detailed information regarding the collection site and collecting procedures must be provided when applying for certification. Photos and herbarium specimens may be required. Germplasm accessions acquired within established protocols of recognized public agencies, however, are normally eligible to enter the certification process as stock seed without routine certification agency site inspections. For contract grow-outs, however, this collection site information must be provided to the grower to enable certification.
Collection Timing
Indeterminate flowering and seed production suggests a broad collection window for spreading fleabane. The Seeds of Success program made nine spreading fleabane seed collections between 2003 and 2018 (USDI BLM SOS 2021). Seed was harvested in Montana, Utah, Colorado, California, and Arizona. The earliest collection was made on May 24, 2007, at a 4,700-ft (1,433 m) elevation site in Maricopa County, Arizona. The latest harvest was September 15, 2010, at a 7,000-ft (2,134 m) elevation site in Montezuma County, Colorado. At two sites, there were additional harvests made 5 to 13 days after the first (USDI BLM SOS 2021).
Collection Methods
Spreading fleabane seed is collected by stripping or clipping the seed heads (Fig. 8) or clipping flowering (Fig. 9) stems over a breathable container. The seed is tiny and easily wind dispersed.

Figure 8. Harvest of spreading fleabane seed made by Seeds of Success collection crews on the Colorado Plateau. Photo: USDI, BLM CP1, SOS.

Figure 9. Collection of spreading fleabane stems clipped from plants in California. Photo: USDI, BLM CA320, SOS.
Several collection guidelines and methods should be followed to maximize the genetic diversity of wildland collections: 1) collect seed from a minimum of 50 randomly selected plants; 2) collect from widely separated individuals throughout a population without favoring the most robust or avoiding small stature plants; and 3) collect from all microsites including habitat edges (Basey et al. 2015). General collecting recommendations and guidelines are provided in online manuals (e.g., ENSCONET 2009; USDI BLM SOS 2021).
It is critical that wildland seed collection does not impact the sustainability of native plant populations. Collectors should take no more than 20% of the viable seed available at the time of harvest (USDI BLM SOS 2021). Additionally, care must be taken to avoid the inadvertent collection of weedy species, particularly those that produce seeds similar in shape and size to those of spreading fleabane.
Post-Collection Management
Although not specifically discussed in the literature, post-collection management of spreading fleabane seed likely requires attention like that of most wildland seed. Seed needs to be dried thoroughly before storing, and if insects are suspected, insecticidal strips can be used or seed can be stored at low temperatures to prevent substantial loss to insect damage.
Seed Cleaning
The Bend Seed Extractory used the following procedure to clean a very small lot (0.035 lb) of spreading fleabane seed. Small seed lots of very small-sized seeds are challenging to clean and require the use of specialized equipment.
Seed was first processed through a Hoffman Hand Deawner/Debearder to remove the seed from the flowers. Next seed was processed through a Laboratory Test Sieve with a mesh size of 30 to remove other non-seed material. Seed was then air screened to remove any remaining inert material using an office Clipper with a 40 × 40 wire top screen, blank bottom screen, and low speeds and air (Barner 2009).
Seed Storage
Spreading fleabane seed is orthodox. After 2 months of storage at 15% relative humidity and -4 °F (-20 °C), seed viability of spreading fleabane was 98%. The initial viability was unknown (SER, INSR, RBGK 2023).
Seed Testing
Moore (1985) provided the following unofficial tetrazolium (TZ) test of spreading fleabane seed viability. Seed was moistened 6 to 8 hours, cut longitudinally with a complete cut through the midsection of the distal half, and the distal end of the seed removed. The cut seed was stained for 6 to 24 hours and considered viable if stained completely or all but the distal third was stained (Moore 1985).
There is no AOSA rule for testing germination of spreading fleabane seed, but there is one for aspen fleabane, which may provide a good starting point. The amount of seed needed for purity testing is also provided for aspen fleabane (AOSA 2016).
Germination Biology
Studies suggest that spreading fleabane seed is nondormant and germinates without pretreatment, but these studies lacked details about where the seed was collected and stored prior to testing.
Laboratory tests reported 96 to 98% germination at incubation temperatures of 59 to 68 °F (15-20 °C) and 89% at 50 °F (10 °C) incubation. Seed was incubated on 1% agar and exposed to an 8/16 light:dark cycle [SER, INSR, RBGK 2023].
In a study of vegetation dynamics in chaparral vegetation, researchers found that germination of spreading fleabane was completely inhibited by leaf leachate of chamise. Germination of spreading fleabane was up to 70% when seed was moistened with untreated water. Germination of both seed treatments was evaluated after 72 hours at 73 °F (23 °C) incubation. Additional support for allelopathic properties of chamise included finding spreading fleabane only in artificially created openings or burned areas within chaparral communities (Christensen and Muller 1975).
Wildland Seed Yield And Quality
Post-cleaning seed yield and quality of seed lots collected in the Intermountain region are provided in Table 2 (USDA FS BSE 2021). The results indicate that spreading fleabane seed is small and can generally be cleaned to high levels of purity.
Table 2. Seed yield and quality of spreading fleabane seed lots collected in the Intermountain region, cleaned and evaluated by the Bend Seed Extractory. Viability was tested by the Oregon State Seed Laboratory or the USDA Forest Service National Seed Laboratory (USDA FS BSE 2021).
| Seed lot characteristic | Mean | Range | Samples (no.) |
| Bulk weight (lbs) | 0.16 | 0.035-0.48 | 8 |
| Clean weight (lbs) | 0.01 | 0.001-0.034 | 8 |
| Clean-out ratio | 0.09 | 0.017-0.171 | 8 |
| Purity (%) | 88 | 78-99 | 8 |
| Fill (%)¹ | 92 | 80-99 | 8 |
| Viability (%)² | 80 | 45-95 | 8 |
| Seeds/lb | 9,640,285 | 1,905,882-15,120,000 | 8 |
| Pure live seeds/lb | 6,440,806 | 1,412,259-10,039,680 | 8 |
¹ 100 seed X-ray test
² Tetrazolium chloride test
Marketing Standards
Acceptable seed purity, viability, and germination specifications vary with revegetation plans. Purity needs are highest for precision seeding equipment used in nurseries, while some rangeland seeding equipment handles less clean seed quite well.
Agricultural Seed Production
There were no reports of growing spreading fleabane in an agricultural setting for native seed production.
Agricultural Seed Certification
In order to minimize genetic changes in specific accessions of native species when increased in cultivated fields, it is essential to track the geographic source and prevent inadvertent hybridization or selection pressure. This is accomplished by following third party seed certification protocols for Pre-Variety Germplasm (PVG) as established by the Association of Official Seed Certification Agencies (AOSCA). AOSCA members in the U.S., Canada, and other countries administer PVG requirements and standards that track the source and generation of planting stock. Field and cleaning facility inspections then monitor stand establishment, proper isolation distances, control of prohibited weeds, seed harvesting, cleaning, sampling, testing, and labeling for commercial sales (UCIA 2015; Young et al. 2020).
Seed growers apply for certification of their production fields prior to planting and plant only certified stock seed of an allowed generation (usually less than four). The systematic and sequential tracking through the certification process requires preplanning, knowing state regulations and deadlines, and is most smoothly navigated by working closely with state certification agency personnel. See the Wildland Seed Certification section for more information on stock seed sourcing.
Nursery Practice
Production of nursery stock of spreading fleabane was not described in the reviewed literature.
Wildland Seeding And Planting
Use of spreading fleabane in wildland restoration or revegetation was not reported in the reviewed literature.
Acknowledgements
Funding for Western Forbs: Biology, Ecology, and Use in Restoration was provided by the USDI BLM Great Basin Native Plant Materials Ecoregional Program through the Great Basin Fire Science Exchange. Great thanks to the chapter reviewers: Judy Perkins, Mojave Desert Ecoregional Native Plant Coordinator, Bureau of Land Management and Shaun Bushman, Plant Research Geneticist, Agricultural Research Service.
This research was supported in part by the USDA Forest Service, Rocky Mountain Research Station. The findings and conclusions in this publication are those of the authors and should not be construed to represent any official USDA or U.S. Government determination or policy.
Literature Cited
Abella, S.R. 2009a. Tree canopy types constrain plant distributions in ponderosa pine-Gambel oak forests, northern Arizona. Res. Note. RMRS-RN-39. Fort Collins, CO: U.S. Department of Agriculture, U.S. Forest Service, Rocky Mountain Research Station. 7 p.
Abella, S.R. 2009b. Smoke-cued emergence in plant species of ponderosa pine forests: Contrasting greenhouse and field results. Fire Ecology. 5(1): 22-37.
Abella, S.R.; Covington, W.W. 2006. Vegetation-environment relationships and ecological species groups of an Arizona Pinus ponderosa landscape, USA. Plant Ecology. 185(2): 255-268.
Abella, S.R.; Springer, J.D.; Covington, W.W. 2007. Seed banks of an Arizona Pinus ponderosa landscape: Responses to environmental gradients and fire cues. Canadian Journal of Forest Research. 37(3): 552-567.
Arathi, H.S.; Hardin, J. 2021. Pollinator-friendly flora in rangelands following control of cheatgrass (Bromus tectorum): A case study. Invasive Plant Science and Management. 14(4): 270-277.
Association of Official Seed Analysts [AOSA]. 2016. AOSA rules for testing seeds. Vol. 1. Principles and procedures. Washington, DC: Association of Official Seed Analysts.
Barner, J. 2009. Propagation protocol for production of propagules (seeds) Erigeron divergens (Torr. & A. Gray). Native Plant Network. U.S. Department of Agriculture, Forest Service, National Center for Reforestation, Nurseries, and Genetic Resources.
Basey, A.C.; Fant, J.B.; Kramer, A.T. 2015. Producing native plant materials for restoration: 10 rules to collect and maintain genetic diversity. Native Plants Journal. 16(1): 37-53.
Bock, J.H.; Bock, C.E. 1992. Short-term reductions in plant densities following prescribed fire in an ungrazed semidesert shrub-grassland. The Southwestern Naturalist. 37(1): 49-53.
Bower, A.D.; St. Clair, J.B.; Erickson, V. 2014. Generalized provisional seed zones for native plants. Ecological Applications. 24(5): 913-919.
Bowers, J.E. 1980. Flora of Organ Pipe Cactus National Monument. Journal of the Arizona-Nevada Academy of Science. 15(2): 33-47.
Bruner, W.E. 1931. The vegetation of Oklahoma. Ecological Monographs. 1(2): 100-188.
Buechner, H.K. 1950. Life history, ecology, and range use of the pronghorn antelope in Trans-Pecos Texas. American Midland Naturalist. 43(2): 257-354.
Capinera, J.L. 2001. Order Orthoptera – Grasshoppers and crickets. In: Handbook of vegetable pests. San Diego, CA: Academic Press: 511-534.
Capinera, J.L. 2020. Chapter 11: Order Orthoptera – Grasshoppers, crickets, and katydids. In: Handbook of vegetable pests (Second Edition). San Diego, CA: Academic Press: 555-579.
Carvajal-Acosta, A.N.; Abella, S.R.; Thompson, D.B. 2015. Initial vegetation response to fuel mastication treatments in rare butterfly habitat of the Spring Mountains, Nevada. Journal of the Arizona-Nevada Academy of Science. 46(1): 6-17.
Christensen, N.L.; Muller, C.H. 1975. Effects of fire on factors controlling plant growth in Adenostoma chaparral. Ecological Monographs. 45(1): 29-55.
Concilio, A.L.; Nippert, J.B.; Ehrenfeucht, S.; Cherwin, K.; Seastedt, T.R. 2016. Imposing antecedent global change conditions rapidly alters plant community composition in a mixed-grass prairie. Oecologia. 182(3): 899-911.
Consortium of Pacific Northwest Herbaria [CPNWH]. 2020. Seattle, WA: University of Washington Herbarium, Burke Museum of Natural History and Culture. pnwherbaria.org/
Crawford, J.A.; Wahren, C.H.A.; Kyle, S.; Moir, W.H. 2001. Responses of exotic plant species to fires in Pinus ponderosa forests in northern Arizona. Journal of Vegetation Science. 12(2): 261-268.
Crimmins, T.M.; Crimmins, M.A.; Bertelsen, C.D. 2010. Complex responses to climate drivers in onset of spring flowering across a semi-arid elevation gradient. Journal of Ecology. 98(5): 1042-1051.
Cronquist, A. 1947. Systematic treatment of the species: [Erigeron compositus–Erigeron acris]. Brittonia. 6(2): 242-300.
Cronquist, A. 1994. Volume five: Asterales. In: Cronquist, A.; Holmgren, A.H.; Holmgren, N.H.; Reveal, J.L.; Holmgren, P.K., eds. Intermountain flora: Vascular plants of the Intermountain West, U.S.A. Bronx, NY: The New York Botanic Garden. 496 p.
Discover Life. 2024. Sam Houston State University. Available: https://www.discoverlife.org/
Drezner, T.D.; Fall, P.L. 2002. Effects of inter-annual precipitation patterns on plant cover according to dispersal mechanisms along a riparian corridor in the Sonoran Desert, U.S.A. Journal of the Arizona-Nevada Academy of Science. 34(2): 70-80.
Elmore, F.H. 1944. Ethnobotany of the Navajo: A monograph of the University of New Mexico and the School of American Research No. 8. Santa Fe, NM: University of New Mexico. 136 p.
European Native Seed Conservation Network [ENSCONET]. 2009. ENSCONET seed collecting manual for wild species. Edition 1: 32 p.
Evenhuis, N.L. 1989. A new species of Mythicomyia from central Arizona (Diptera: Bombyliidae). The Southwestern Naturalist. 34(4): 527-530.
Fagan, D. 1998. Canyon country wildflowers. A Falcon Guide. Helena, MT: Falcon Publishing Company. 147 p.
Flake, L.D. 1973. Food habits of four species of rodents on a short-grass prairie in Colorado. Journal of Mammalogy. 54(3): 636-647.
Flinders, J.T.; Hansen, R.M. 1972. Diets and habitats of jackrabbits in northeastern Colorado. Range Science Department Science Series No. 12. Fort Collins, CO : Colorado State University. 29 p.
Gardner, J.L.; Hubbell, D.S. 1934. Some vegetational responses after eight years of protection from grazing. Ecology. 24(3): 409-410.
Goodwin, J.G., Jr.; Hungerford, C.R. 1979. Rodent population densities and food habits in Arizona ponderosa pine forests. Research Paper RM 214. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Forest and Range Experiment Station. 12 p.
Grand Prismatic Seed (2024). Online seed sales. Fruit Heights, UT. Available: https://www.grandprismaticseed.com/archived/spreading-fleabane
Guo, Q. 1996. Effects of bannertail kangaroo rat mounds on small-scale plant community structure. Oecologia. 106(2): 247-256.
Guo, Q.; Brown, J.H.; Valone, T.J. 2002. Long-term dynamics of winter and summer annual communities in the Chihuahuan Desert. Journal of Vegetation Science. 13(4): 575-584.
Hannon, L.E.; Sisk, T.D. 2009. Hedgerows in an agri-natural landscape: Potential habitat value for native bees. Biological Conservation. 142(10): 2140-2154.
Hayward, C.L. 1948. Biotic communities of the Wasatch chaparral, Utah. Ecological Monographs. 18(4): 473-506.
Herron-Sweet, C.R.; Lehnhoff, E.A.; Burkle, L.A.; Littlefield, J.L.; Mangold, J.M. 2016. Temporal‐ and density‐dependent impacts of an invasive plant on pollinators and pollination services to a native plant. Ecosphere. 7(2): e01233.
Hickman, J.C., ed. 1993. The Jepson manual: Higher plants of California. Berkeley, CA: University of California Press. 1400 p.
Hitchcock, C.L.; Cronquist, A. 2018. Flora of the Pacific Northwest: An illustrated manual. Second Ed. Giblin, D.E.; Legler, B.S.; Zika, P.F.; Olmstead, R.G., eds. Seattle, WA: University of Washington Press. 882 p.
Humphrey, R.R. 1955. Forage production on Arizona ranges: IV. Coconino, Navajo, Apache Counties: A study in range condition. Bulletin No. 266. Tucson, AZ: University of Arizona, Agricultural Experiment Station. 84 p.
James, D.G.; Nunnallee, D. 2011. Life histories of Cascadia butterflies. Corvallis, OR: Oregon State University Press. 447 p.
Keil, D.J., Pinkava, D.J. 1976. Chromosome counts and taxonomic notes for Compositae from the United States and Mexico. American Journal of Botany. 63(10): 1393-1403.
Korb, J.E.; Johnson, N.C.; Covington, W.W. 2004. Slash pile burning effects on soil biotic and chemical properties and plant establishment: Recommendations for amelioration. Restoration Ecology. 12(1): 52-62.
Korb, J.E.; Stoddard, M.T.; Huffman, D.W. 2020. Effectiveness of restoration treatments for reducing fuels and increasing understory diversity in shrubby mixed-conifer forests of the southern Rocky Mountains, USA. Forests. 11(5): 508.
Krausman, P.R. 1978. Forage relationships between two deer species in Big Bend National Park, Texas. Journal of Wildlife Management. 42(1): 101-107.
Krausman, P.R.; Kuenzi, A.J.; Etchberger, R.C.; Rautenstrauch, K.R.; Ordway, L.L.; Hervert, J.J. 1997. Diets of desert mule deer. Journal of Range Management. 50(5): 513-522.
Lady Bird Johnson Wildflower Center [LBJWC]. 2022. Erigeron divergens (Torrey & A. Gray). Native Plant Database. Austin, TX: Lady Bird Johnson Wildflower Center. https://www.wildflower.org/plants-main [Accessed 2024 May 4].
Lambert, S. 2005. Guidebook to the seeds of native and non-native grasses, forbs and shrubs of the Great Basin. Boise, ID: U.S. Department of the Interior, Bureau of Land Management, Idaho State Office. 136 p.
Lance, R.F.; Bailey, P.; Lindsay, D.L.; Cobb, N.S. 2017. Precipitation and the robustness of a plant and flower-visiting insect network in a xeric ecosystem. Journal of Arid Environments. 144(2017): 48-59.
Lesica, P. 2012. Manual of Montana vascular plants. Fort Worth, TX: Botanical Research Institute of Texas (BRIT) Press. 779 p.
Lindsey, A.A. 1951. Vegetation and habitats in a southwestern volcanic area. Ecological Monographs. 21(3): 227-253.
Love, A. 1974. IOPB chromosome number reports. XLV. Taxon. 23(4): 619-624.
MacKinnon, A.; Kershaw, L.; Arnason, J.; Owen, P.; Karst, A.; Hamersley Chambers, F. 2014. Edible and medicinal plants of Canada. Edmonton, Alberta: Partners Publishing and Lone Pine Publishing. 448 p.
McArthur, E.D.; Blauer, A.C.; Sanderson, S.C. 1988. Mule deer-induced mortality of mountain big sagebrush. Journal of Range Management. 41(2): 114-117.
McCulloch, C.Y. 1973. Part I: Seasonal diets of mule and white-tailed deer. In: Deer nutrition in Arizona chaparral and desert habitats. Special Report No. 3: Federal Aid in Wildlife Restoration Act Project W-78-R. Phoenix, AZ: Arizona Game and Fish Department, Research Division: 1-37.
Metlen, K.L.; Dodson, E.K.; Erich, K.; Fiedler, C.E. 2006. Research Project Summary: Vegetation response to restoration treatments in ponderosa pine-Douglas-fir forests of western Montana. https://www.fs.fed.us/database/feis
Moore, R.P. 1985. Handbook on tetrazolium testing. Zurich, Switzerland: International Seed Testing Association. 99 p.
Mower, K.J.; Smith, H.D. 1989. Diet similarity between elk and deer in Utah. The Great Basin Naturalist. 49(4): 552-555.
Munz, P.A.; Keck, D.D. 1973. A California flora and supplement. Berkeley, CA: University of California Press. 1905 p.
Nelson, G.H.; Westcott, R.L. 1995. Three new species of Acmaeodera Eschscholtz (Coleoptera: Buprestidae) from the United States and Mexico. The Coleopterists Bulletin. 49(1): 77-87.
Nesom, G.L. 2006. 186. Erigeron. In: Flora of North America Editorial Committee, ed. Flora of North America North of Mexico. Volume 20 Magnoliophyta: Asteridae, part 7: Asteraceae, part 2 Asterales, part 2 (Aster order). New York, NY: Oxford University Press: 256-348.
Nesom, G.L. 2015. Variants of Erigeron divergens (Asteraceae). Phytoneuron. 2015-16: 1-24.
Omernik, J.M. 1987. Ecoregions of the conterminous United States. Map (scale 1:7,500,000). Annals of the Association of American Geographers. 77(1): 118-125.
Pinkava, D.J.; Keil, D.J. 1977. Chromosome counts of Compositae from the United States and Mexico. American Journal of Botany. 64(6): 680-686.
Plant Conservation Alliance [PCA]. 2015. National seed strategy for rehabilitation and restoration 2015-2020. Washington, DC: U.S. Department of the Interior, Bureau of Land Management. 52 p.
Ramaley, R. 1919. Xerophytic grasslands at different altitudes in Colorado. Bulletin of the Torrey Botanical Club. 46(2): 37-52.
Reed, J.F. 1952. The vegetation of the Jackson Hole Wildlife Park, Wyoming. The American Midland Naturalist. 48(3): 700-729.
Richardson, B.; Kilkenny, F.; St. Clair, B.; Stevenson-Molnar, N. 2020. Climate Smart Restoration Tool. https://climaterestorationtool.org/csrt/
Riser, J.P.; Meyers, S.C. 2020. Erigeron (Fleabane, daisy). In: Meyers, S.C.; Jaster, T.; Mitchell, K.E.; Harvey, T.; Hardison, L.K., eds. Flora of Oregon, Volume 2: Dicots A-F. Fort Worth, TX: Botanical Research Institute of Texas (BRIT) Press: 249-262.
Robinson, G.S.; Ackery, P.R.; Kitching, I.; Beccaloni, G.W.; Hernández, L.M. 2023. HOSTS: A Database of the World’s Lepidopteran Hostplants [Data set]. Natural History Museum. https://doi.org/10.5519/havt50xw
Rodriguez-Chavez, J.L.; Egas, V.; Linres, E.; Bye, R.; Hernandez, T.; Espinosa-Garcia, F.J.; Delgado, G. 2017. Mexican arnica (Heterotheca inuloides Cass. Asteraceae: Astereae): Ethnomedical uses, chemical constituents and biological properties. Journal of Ethnopharmacology. 195(2017): 39-63.
Samson, D.A.; Philippi, T.E.; Davidson, D.W. 1992. Granivory and competition as determinants of annual plant diversity in the Chihuahuan Desert. Oikos. 65(1): 61-80.
Schwartz, C.C.; Nagy, J.G. 1976. Pronghorn diets relative to forage availability in northeastern Colorado. The Journal of Wildlife Management. 40(3): 469-478.
SEINet – Regional Networks of North American Herbaria Steering Committee [SEINet]. 2020. SEINet Regional Networks of North American Herbaria. https://Symbiota.org/docs/seinet
Shive, K.L.; Kuenzi, A.M.; Sieg, C.H.; Fule, P.Z. 2013. Pre-fire fuel reduction treatments influence plant communities and exotic species 9 years after a large wildfire. Applied Vegetation Science. 16(3): 457-469.
Shryock, D.F.; DeFalco; L.A.; Esque, T.C. 2018. Spatial decision-support tools to guide restoration and seed-sourcing in the Desert Southwest. Ecosphere. 9(10): p.e02453.
Smiley, F.J. 1915. The alpine and subalpine vegetation of the Lake Tahoe region. Botanical Gazette. 59(4): 265-286.
Society for Ecological Restoration; International Network for Seed Based Restoration; Royal Botanic Gardens [SER, INSR, RBGK]. 2023. Seed Information Database (SID). Available from: https://ser-sid.org/
Spellenberg, R. 2001. National Audubon Society field guide to North American wildflowers: Western region, revised edition. New York, NY: Alfred A. Knopf, Inc. 862 p.
Spellenberg, R. 2012. Sonoran Desert wildflowers: A guide to common plants. Helena, MT: Falcon Guides. 245 p.
Springer, J.D. 1999. Soil seed bank in southwestern ponderosa pine: Implications for ecological restoration. Flagstaff, AZ: Northern Arizona University. Thesis. 101 p.
Springer, J.D.; Huffman, D.W.; Stoddard, M.T.; Sánchez Meador, A.J.; Waltz, A.E.M. 2018. Plant community dynamics following hazardous fuel treatments and mega-wildfire in a warm-dry mixed-conifer forest of the USA. Forest Ecology and Management. 429(2018): 278-286.
Stevens, L.E.; Schmidt, J.C.; Ayers, T.J.; Brown, B.T. 1995. Flow regulation, geomorphology, and Colorado River marsh development in the Grand Canyon, Arizona. Ecological Applications. 5(4): 1025-1039.
Stoddard, M.T.; McGlone, C.M.; Fule, P.Z.; Laughlin, D.C.; Daniels, M.L. 2011. Native plants dominate understory vegetation following ponderosa pine forest restoration treatments. Western North American Naturalist. 71(2): 206-214.
Strother, J.L. 1983. More chromosome studies in Compositae. American Journal of Botany. 70(8): 1217-1224.
Taylor, R.J. 1992. Sagebrush country: A wildflower sanctuary. Missoula, MT: Mountain Press Publishing Company. 211 p.
Trappe, J.M.; Manassah, J.T.; Briskey, E.J. 1981. Mycorrhizae and productivity of arid and semiarid rangelands. In: Manassah, J.T.; Briskey, E.J., eds. Advances in food-producing systems for arid and semiarid Lands. San Francisco, CA: Academic Press: 581-599.
Turner, N.J.; Thompson, L.C.; Thompson, M.T.; York, A.Z. 1990. Thompson ethnobotany: Knowledge and usage of plants by the Thompson Indians of British Columbia. Royal British Columbia Museum Memoir No. 3. Victoria, British Columbia, Canada: Royal British Columbia Museum. 335 p.
USDA Forest Service [USDA FS] 1937. Range plant handbook. Washington, DC: U.S. Department of Agriculture, Forest Service. 816 p.
USDA Forest Service, Bend Seed Extractory [USDA FS BSE]. 2021. Nursery Management Information System Version 4.1.11. Local Source Report 34-Source Received. Bend, OR: U.S. Department of Agriculture, Forest Service, Bend Seed Extractory.
USDA Forest Service, Western Wildland Environmental Threat Assessment Center [USFS WWETAC]. 2017. TRM Seed Zone Applications. Prineville, OR: U.S. Department of Agriculture, Forest Service, Western Wildland Environmental Threat Assessment Center. https://research.fs.usda.gov/pnw/products/dataandtools/seed-zone-webmap
USDA Natural Resources Conservation Service [USDA NRCS]. 2024. The PLANTS Database. Greensboro, NC: U.S. Department of Agriculture, Natural Resources Conservation Service, National Plant Data Team. https://plants.sc.egov.usda.gov/
USDI Bureau of Land Management, Seeds of Success [USDI BLM SOS]. 2020. Seeds of Success collection data. Washington, DC: U.S. Department of the Interior, Bureau of Land Management, Plant Conservation and Restoration Program.
USDI Bureau of Land Management, Seeds of Success [USDI BLM SOS]. 2023. Bureau of Land Management technical protocol for the collection, study, and conservation of seeds from native plant species for Seeds of Success. Washington, DC: U.S. Department of the Interior, Bureau of Land Management, Plant Conservation and Restoration Program. 45 p.
USDI Environmental Protection Agency [USDI EPA]. 2018. Ecoregions. Washington, DC: U.S. Environmental Protection Agency. https://www.epa.gov/eco-research/ecoregions
USDI Geological Survey [USDI USGS]. 2020. Biodiversity Information Serving Our Nation (BISON). U.S. Geological Survey. https://www.gbif.us/
Utah Crop Improvement Association [UCIA]. 2015. How to be a seed connoisseur. Logan, UT: UCIA, Utah Department of Agriculture and Food, Utah State University and Utah State Seed Laboratory. 16 p.
Varva, M.; Rice, R.W.; Hansen, R.M. 1978. A comparison of esophageal fistula and fecal material to determine steer diets. Journal of Range Management. 31(1): 11-13.
Vestal, P.A. 1952. Ethnobotany of the Ramah Navaho. Reports of the Ramah Project: No. 4. Papers of the Peabody Museum of American Archeology and Ethnology. Cambridge, MA: Harvard University. 40(4): 1-94.
Vestal, P.A.; Schultes, R.E. 1939. The economic botany of the Kiowa Indians as it related to the history of the tribe. Cambridge, MA: Botanical Museum. 110 p.
Wagner, W.L.; Martin, W.C.; Aldon, E.F. 1978. Natural succession on strip-mined lands in northwestern New Mexico. Reclamation Review. 1: 67-73.
Welsh, S.L.; Atwood, N.D.; Goodrich, S.; Higgins, L.C., eds. 2015. A Utah Flora. Fifth Edition, revised. Provo, UT: Brigham Young University. 990 p.
Wyman, L.C.; Harris, S.K. 1951. The ethnobotany of the Kayenta Navaho: An analysis of the John and Louisa Wetherill ethnobotanical collection. Publications in Biology. No. 5 Albuquerque, NM: University of New Mexico Press. 66 p.
Young, S.A.; Schrumpf, B.; Bouck, M.; Moore, M. 2020. How the Association of Official Seed Certifying Agencies (AOSCA) tracks wildland sourced seed and other plant propagating materials. Logan, UT: Utah State University, Utah Agricultural Experiment Station. 36 p.
How to Cite
Gucker, Corey L.; Shaw, Nancy L. 2024. Spreading fleabane (Erigeron divergens). In: Gucker, C.L.; Shaw, N.L., eds. Western forbs: Biology, ecology, and use in restoration. Reno, NV: Great Basin Fire Science Exchange. Online: https://westernforbs.org/species/spreading-fleabane-erigeron-divergens/