Authorship
Gucker, Corey; Shaw, Nancy
Publication Date
June 2025
Nomenclature
Utah sweetvetch (Hedysarum boreale) Nutt. belongs to the Multicaulia section of the Hedysareae tribe in the Fabaceae or pea family (Duan et al. 2015; USDA NRCS 2025).
Family
Fabaceae – Pea family
Genus
Hedysarum
Species
boreale
NRCS Plant Code
HEBO (USDA NRCS 2025)
Subtaxa
The Flora of North America recognizes two subspecies: Hedysarum boreale subsp. boreale Nutt. and H. b. subsp. mackenziei (Richardson) S.L. Welsh (Ross 2020).
Synonyms
Hedysarum cinerascens Rydb., H. mackenziei Richarson, H. pabulare A. Nelson, H. utahense Nutt., H. boreale var. cinerascens (Rydb.) Rollins, H. b. var. gremiale (Rollins) Northstrom & S.L. Welsh, H. b. var. obovatum Rollins, H. b. var. pabulare (A. Nelson) Dorn, H. b. var. rivulare (L.O. Williams) Northstrom, H. b. var. typicum Rollins, and H. b. var. utahense (Rydb.) Rollins (Ross 2020; ITIS 2024; USDA NRCS 2025).
Common Names
Utah sweetvetch, boreale sweetvetch, chain-pod, northern sweet broom, northern sweetvetch, and Rocky Mountain sweetvetch (Rosales 2014; ITIS 2024; USDA NRCS 2025).
Chromosome Number
The chromosome number is 2n = 16 (Löve 1982; Welsh et al. 2015).
Hybridization
There were no descriptions of hybrids or hybridization found for this species in the reviewed literature.
Distribution
Utah sweetvetch has a broad distribution from ranging from Alaska, across most of Canada, and south through the western United States to Nevada, Arizona, New Mexico, and Texas. It is particularly common in the Rocky Mountains and other mountainous regions of the western United States (Hitchcock and Cronquist 2018; Ross 2020). Subspecies mackenzii occupies only the northern range of the species, occurring from Alaska across Canada and south to Washington and Oregon (Hitchcock and Cronquist 2018). Subspecies boreale occurs from British Columbia to Manitoba and south, mainly in the Rocky Mountain states, but also in northeastern Oregon, the Dakotas, Nevada, Arizona, New Mexico, and Texas (Hitchcock and Cronquist 2018; Ross 2020).
Habitat And Plant Associations
As its broad distribution suggests, Utah sweetvetch is adapted to a diversity of site conditions and grows in habitats ranging from desert shrublands to tundra (Barneby 1989; Ford et al. 1989; Welsh et al. 2015). In the United States, Utah sweetvetch is common at low to mid-elevations receiving at least 10 in (250 mm) of annual precipitation (Ford et al. 1989; Ogle et al. 2003). It grows in grasslands; sagebrush (Artemisia spp.), oak brush (Quercus spp.), and mountain brush shrublands; pinyon-juniper (Pinus–Juniperus spp.) and quaking aspen (Populus tremuloides) woodlands; ponderosa pine (Pinus ponderosa) forests; alpine communities; and riparian sites (Figs. 1 and 2) (Plummer et al. 1968; Goodrich and Neese 1986; Barneby 1989; Lesica 2012; Welsh et al. 2015). It favors dry, open to lightly shaded sites and is rarely found growing in dense vegetation with high levels of shading or plant competition (USDA FS 1937; Redente 1980). In Utah rangelands it is commonly associated with western wheatgrass (Pascopyrum smithii), Salina wildrye (Leymus salinus), juniper, sagebrush, and globemallow (Sphaeralcea spp.) (Parker 1980; Rumbaugh and Johnson 1984).
Figure 1. Utah sweetvetch growing in a drainage area in a big sagebrush shrubland in Beaverhead, Montana. Photo: M. Lavin, Montana State University (MSU).
Elevation
Throughout its range, Utah sweetvetch occurs from 2,950 ft (900 m) to 10,000 ft (3,000 m) (USDA FS 1937; Barneby 1989; Welsh et al. 2015; Ross 2020).
Soils
Utah sweetvetch grows in moderately acidic to alkaline, fine to coarse-textured, rocky or gravelly soils ranging from sands to heavy clays (Fig. 2) (Taylor 1992; Jensen et al. 1999; Ogle et al. 2003; Rosales 2014; Ross 2020) but is most common in well-drained sandy to silty clay loams (Stevens et al. 1996). Plants have high drought tolerance, moderate salinity tolerance, and low fertility requirements (Mee et al. 2003; LBJWC 2013; Rosales 2014). Soils evaluated at 18 Utah sweetvetch sites in the Piceance Basin in northwestern Colorado were loams to sandy loams. The gravel content of these soils ranged from 16 to 58%, pH from 7 to 8.6, organic matter from 1.3 to 4.3%, nitrate from 6 to 16 ppm, and plant available phosphorus from 1.5 to 4 ppm (Redente 1980).

Figure 2. Utah sweetvetch growing in rocky soils along a small creek in Alaska. Photo: U.S. Department of the Interior, Bureau of Land Management (DOI, BLM) AK930 Seeds of Success (SOS).
In the United States, Utah sweetvetch is associated with soils derived from sedimentary and volcanic rock. Plants are locally abundant on shale exposures on mesas in the southern Rocky Mountains (Weber 1976) and in alpine habitats on calcareous substrates of limestone parent material in Montana (Bamberg and Major 1968). In Utah, it grows in light sandy soils with shadscale (Atriplex confertifolia) and fourwing saltbush (A. canescens) and in heavy-textured soils where greasewood (Sarcobatus vermiculatus) comes into the shadscale-saltbush community. Plants are associated with neutral to acidic soils from volcanic parent materials and alkaline soils from sedimentary parent materials (Plummer et al. 1968).
Several sources indicate that Utah sweetvetch has some tolerance of metals and salt. Utah sweetvetch grows in salt desert shrublands with moderate soil salinity, alkalinity, and some exchangeable sodium (Wasser and Shoemaker 1982). In Arizona, Utah sweetvetch is listed as a restoration plant for sites with metals or salts (EcoRestore AZ 2024). In a controlled study, Utah sweetvetch seedling mortality averaged 47% with exposure to 4 g/l sodium chloride, but biomass production of surviving seedlings was not significantly different from controls (Boukheloua 1990).
Utah sweetvetch is drought tolerant based on observational and experimental studies. Rouse and Kershaw (1973) evaluated plant communities along an elevation gradient from shoreline to uplands near Hudson Bay in Ontario, Canada. Cover of Utah sweetvetch was 5% at the youngest, wind-scoured, sparsely vegetated shoreline site, 3% at the next youngest site, 2% at a poorly drained lowland site, and 9% at a more upland site. Volumetric soil moisture at 0 to 28 in (70 cm) depths at these four sites was 8%, 21%, 38%, and 26%, respectively (Rouse and Kershaw 1973). At the Mancos Shale Barrens in Dolores, Colorado, Utah sweetvetch grew in shallow silt-clay soils (4 to 6 in [10-15 cm] deep) where soil moisture averaged 9% (Silva and Ayers 2016).
Utah sweetvetch seedlings from 11 populations exhibited drought tolerance in greenhouse experiments (Ford et al. 1989; Johnson et al. 1989). Seed collected from southwestern Montana, southern Idaho, Utah, and eastern Colorado locations were established with watering levels at near field capacity for 35 days. Seedlings were then grown for 38 days with abundant (3.4 in [8.7 cm]), adequate (2 in [5.3 cm]), and limited (0.2 in [0.4 cm]) total water supplied. Seedling mortality was minimal with the abundant and adequate water level treatments, but 30% with limited water. Shoot and root mass as well as the mass of root nodules generally decreased with decreasing water availability. The proportion of root mass to total plant mass increased as less water was applied. There were significant tolerance and growth differences among the populations. One produced its heaviest shoots with abundant water, another with limited water, and yet another with relatively constant shoot weights regardless of irrigation amounts. Total root length also differed among populations with some producing their longest roots with abundant and adequate water treatments and another with limited water (Ford et al. 1989; Johnson et al. 1989). See Nursery Production for a description of seedling growth conditions.
Description
Utah sweetvetch is a perennial, tap-rooted legume with a stout, superficial or shallowly buried woody caudex and a bushy appearance (Redente 1980; Barneby 1989; Taylor 1992; Mee et al. 2003; Welsh et al. 2015; Ross 2020). Taproots are stout, woody, and deep; crowns are robust (USDA FS 1937; Jensen et al. 1999; Ross 2020). Plants are highly variable with mostly erect, 4 to 28 in (10-70 cm) tall flowering stems (Fig. 3). Stems are simple or few-branched and somewhat spreading at the base, or they may extend laterally near the ground before curving upward (USDA FS 1937; Barneby 1989; Welsh et al. 2015). Herbage ranges from green and almost glabrous to gray and densely hairy (Figs. 3 and 4) (Barneby 1989; Lesica 2012).

Figure 3. Utah sweetvetch plant with bright green foliage near Butte, Montana. Photo: M. Lavin, Montana State University (MSU).
Plants produce long carrot-like taproots up to several feet in length (Plummer et al. 1968). Roots branching laterally from the taproot spread a few centimeters from the main taproot before extending down in paths that follow the taproot (Redente 1980). Native plant gardeners suggest taproots need a minimum soil depth of 18 in (46 cm) (Mee et al. 2003). In studies of extracted plants, most roots had nitrogen-fixing nodules (Plummer et al. 1968). Some plants produce rhizomes, but clumps forming from rhizomatous types have limited spread (PFAF 2024).

Figure 4. Pubescent foliage gives this Utah sweetvetch plant growing in Utah a gray-green color. Photo: DOI, BLM UT931 SOS.
Leaves are usually opposite and 1 to 6 in (3-15 cm) long. They are odd-pinnately compound with 5 to 15 thick, linear-elliptic to broadly ovate leaflets 1 to 2 cm long (Barneby 1989; Lesica 2012; Welsh et al. 2015; Ross 2020). Leaflets have prominent midribs on the underside and minute brown speckles on the upper side (Barneby 1989). All leaves have two thin, leaflike, brownish bracts at the base (USDA FS 1937).
Utah sweetvetch produces narrow, congested, axillary, pedunculate, and bracteate racemes (Barneby 1989; Taylor 1992; Lesica 2012; Ross 2020). In a pollination study of natural Utah sweetvetch populations in Utah and Wyoming, plants varied from having a few to hundreds of racemes (Swoboda 2007). Individual racemes are narrow, congested, axillary, pedunculate, and bracteate (Barneby 1989; Taylor 1992; Lesica 2012; Ross 2020). The inflorescence axis, including the peduncle, is 2 to 11 in (5-28 cm) long (Barneby 1989; Welsh et al. 2015). Each inflorescence supports 5 to 50 papilionaceous flowers about 0.25 in (0.6 cm) long (Fig. 5) (USDA FS 1937; Belcher 1985; Welsh et al. 2015; Ross 2020). The corolla is rose to dark purple with a 5-toothed calyx that slightly exceeds the tube (Barneby 1989). The keel is abruptly bow-shaped and longer than the wings by up to 3.5 mm (Barneby 989; Welsh et al. 2015; Ross 2020). Flowers have 10 stamens, nine united and one separate (Lesica 2012).

Figure 5. Indeterminately flowering Utah sweetvetch inflorescences with the basal flower opening first. Photo: DOI, BLM UT931 SOS.
Fruits are flattened linear pods (loments) with conspicuous veining that is mostly perpendicular to the axis. They are 4 to 7 mm wide with two to eight one-seeded indehiscent segments that separate at the constrictions (Fig. 6) (USDA FS 1937; Lesica 2012; Welsh et al. 2015; Hitchcock and Cronquist 2018; Ross 2020). The pods are pendulous to spreading and occur in clusters (USDA FS 1937; Plummer et al. 1968; Lesica 2012). Each pod may contain up to eight seeds, but some seeds are normally abortive (Barneby 1989). Seeds are brown and kidney shaped (Rosales 2014).

Figure 6. Utah sweetvetch stems with flowers, fading flowers, and developing pods (loments). Photo: M. Lavin, MSU.
Descriptions of Subspecies. In the Pacific Northwest, Utah sweetvetch subspecies are best distinguished by their inflorescences and flowers (Hitchcock and Cronquist 2018). Subspecies mackenzii is generally glabrous to sparsely strigulose with compact racemes 0.8 to 3 in (2-8 cm) long that produce red-purple flowers (15-26 mm long) that fade to dark purple and loments with 3 to 8 segments. Subspecies boreale is glabrous or pubescent with elongate racemes (3 to 13 in [8-32 cm] long) that produce pink to purple flowers (10-19 mm long) that fade to pink or lavender and loments with 2 to 6 segments (Hitchcock and Cronquist 2018).
Lookalikes. Utah sweetvetch can be easily confused with milkvetch (Astragalus spp.) when not in fruit (Weber 1976; Pavek et al. 2012). It can be distinguished from milkvetch species by the squared tips of its flowers, segmented and veined loments, and leaves with glandular dots that are visible when held to the light (USDA FS 1937).
Belowground Relationships And Interactions
Utah sweetvetch produces root nodules that house nitrogen-fixing bacteria. This relationship improves its growth and the nutrient content of the environment where it grows.
A controlled study of Utah sweetvetch seedlings grown for 12 weeks with and without rhizobia and mycorrhizae fungi found that seedling growth improved with additions of either and both types of amendments (Table 1). Four-day old seedlings were transplanted into pots filled with soil collected from a sagebrush community in the Piceance Basin in northwestern Colorado. Field-collected soil was mixed 2:1 with sand and sterilized. Pots were then inoculated with arbuscular mycorrhiza (Glomus fasciculatus), soil bacteria (Rhizobium spp.), mycorrhiza + soil bacteria, or not inoculated (control). Inoculation with either or both species significantly increased seedling leaf production, height, and above and below ground weight compared to controls. The soil bacteria treatment increased plant nitrogen content, while the mycorrhiza treatment increased the phosphorus content of aboveground foliage (Redente and Reeves 1981).
Table 1. Early seedling growth of Utah sweetvetch in sterilized, field-collected soil with soil bacteria, mycorrhiza, or soil bacteria + mycorrhiza added (Redente and Reeves 1981).
|
Treatment |
Number of leaves |
Height (cm) | Aboveground dry weight (g) | Belowground dry weight (g) |
Root:shoot ratio |
| Control |
42a1 |
19.9a | 5.1a | 5.5a |
1.08a |
| Rhizobium |
69b |
37.5b | 7.5b | 16.6b |
2.21b |
| Glomus |
78bc |
40.2bc | 8.8c | 14.1c |
1.60c |
| Glomus + Rhizobium |
84c |
43.1c | 9.1c | 12.4c |
1.36c |
1Values within a column followed by different letters differ significantly (P ≤ 0.05).
A study of successional plant communities on different aged moraines near Mount Robson glacier in British Columbia found that Utah sweetvetch was the major contributor of nitrogen to moraine soil (Blundon and Dale 1990). Although Utah sweetvetch plants produced the greatest number of nodules (80.3 nodule meristems/plant) in pioneer communities and the least in the oldest communities (6.3 nodule meristems/plant), Utah sweetvetch contributed 72% of the nitrogen to the youngest moraines and 88% to the oldest (Blundon and Dale 1990).
Reproduction
Utah sweetvetch reproduces from seed (USDA FS 1937). Plants of some populations produce rhizomes, a potential form of vegetative reproduction (Shaw 2004). Based on studies of agricultural seed production, Utah sweetvetch typically produces some seed in its second year, and full seed production generally occurs by year 3 (Stevens et al. 1996; Rosales 2014).
Phenology
Utah sweetvetch flowers from May to early August depending on location and weather conditions (Barneby 1989; Ross 2020). First flowers appear in spring, and a second minor flowering can occur if initial fruit set fails (Kitchen 1994). In southeastern Montana, Utah sweetvetch flowering ended by late July in 1975 and 1976. Seed pods began dropping in early August (Eddleman 1977). Plants transition into vegetative dormancy as soil moisture decreases (Kitchen 1994). Phenology is further discussed in the Seed Collection section.
Breeding System
Although Utah sweetvetch is self-compatible, insect visitation is required to produce viable seed (Swoboda and Cane 2012).
Pollination
Pollination of Utah sweetvetch flowers is required for seed production, and bees are the primary pollinators (Tepedino and Stackhouse 1987; McGuire 1993; Kudo and Harder 2005; Swoboda 2007). In cultivated stands of Utah sweetvetch in northern Utah, lower stigmas were receptive for several days beginning at the mature bud stage, and younger flowers (0-24 hrs) were more likely to produce fruit than older flowers (48-72 hrs, P = 0.0006; >72 hrs P = 0.106) (Swoboda 2007). Swoboda (2007) studied controlled and natural pollination of Utah sweetvetch in seven natural populations and three cultivated stands for 2 years in Utah and Wyoming. Self-pollination of 705 individual flowers (autogamy) produced no fruits or seeds. Although cross pollination between different plants (xenogamy) produced fewer fruits than cross pollination between flowers on the same plant (geitonogamy), fruits resulting from xenogamy were twice as likely to contain viable seeds (~75-98%) as those resulting from geitonogamy (~28-48%) (Swoboda 2007; Swoboda and Cane 2012).
In two natural populations of Utah sweetvetch in lower canyon locations of Cache County, Utah, bee pollination resulted in 57% and 46% fruit set (Swoboda 2007; Swoboda and Cane 2012). In 2 years of observations, pollinator visitors included 8 genera and 31 species. Most visitors were of the Apidae and Megachilidae families including bumble bees (Bombus), small mason bees (Hoplitis), leafcutter bees (Megachile), and mason bees (Osmia) (Swoboda 2007).
At the Bonanza Creek Bluff Experimental Forest west of Fairbanks, Alaska, pollination was observed for 40.5 hours at early seral sites in the summer of 1988 (McGuire 1993). Male and female leafcutting bees (M. giliae) made up about 40% of the visits to Utah sweetvetch flowers. Seed set/flower increased with increased visitation by female M. giliae (McGuire 1993).
In montane and subalpine sites in the Kananaskis Valley in Alberta, Utah sweetvetch flowers were exclusively visited by bumble bees (Kudo and Harder 2005), and Utah sweetvetch was one of two legumes visited by overwintered bumble bee queens. Based on observations made in 2001, Utah sweetvetch flowered from late May to late June and flowering lasted 13 to 14 days. Plants averaged 3.4 total open flowers/inflorescence, 1.6 were visited by bumble bees, and seed set/fruit averaged 37% (Kudo and Harder 2005).
Long-tongued bees were the main visitors to Utah sweetvetch growing in a big sagebrush (Artemisia tridentata) shrubland on south and west slopes of Spread Creek Hill, Teton County, Wyoming (~7,200 ft [2,200 m] elevation) (Tepedino and Stackhouse 1987). Pollinator visitors were monitored in a population of several hundred plants. Bees were collected from late June to mid-July at peak flowering (3 time periods/day, between 940 and 2200 for 5 days and a total of 40-man hours). Of the 37 bee species collected, most were Osmia (14), Bombus and Psithyrus (10), and Megachile (4) (Tepedino and Stackhouse 1987).
Ecology
Utah sweetvetch is a moderately long-lived, disturbance-tolerant species that is also an early colonizer in primary and secondary succession. Its stout taproot is deep and tough, making the plant drought resistant, winter hardy, and “renders [it] somewhat resistant to abuse”, and capable of persisting and even increasing on severely grazed rangelands (USDA FS 1937; Hermann 1966; Ogle et al. 2003). Plants are common on roadsides (Fig. 7) (LBJWC 2013) and increases in vegetative and reproductive production are common after fire (Kitchen 1994). Sprouting has also been observed after roots were severely cut (Boukheloua 1990).

Figure 7. Utah sweetvetch plant growing along a gravel road in Bozeman, Montana. Photo: M. Lavin, Montana State University.
Utah sweetvetch has been described as moderately long-lived to long-lived (Northstrom 1969; Kitchen 1994), but only one study attempted to estimate its life span (Treshow and Harper 1974). Life expectancy was projected to be 10 to 20 years when researchers evaluated taproot growth rings, annual mortality, and annual vegetative replacement and survival rates. Over four growing seasons, annual mortality averaged 5% (range: 4.5-6.5%), annual vegetative replacement averaged 3%, and survival averaged 92%. Longevity was evaluated in grass, oak brush, aspen, and conifer forest plots in Utah (Treshow and Harper 1974).
Many studies reported that Utah sweetvetch colonizes sites in the earliest years of succession. Utah sweetvetch was an initial colonizer of Tanana River floodplain habitats in Alaska’s Bonanza Creek Experimental Forest (Hollingsworth et al. 2010). In a successional study on ice moraines of Klutlan Glacier at the Alaska-Yukon Territory border, Utah sweetvetch was an early colonizer, occurring in vegetation where dominant plants were 2 to 30 yrs old (Birks 1980). It also naturally colonized 5 of 16 man-made gravel pads abandoned 10 years previously in arctic Alaska. Soil in the pads had little organic matter or fine material, low water-holding capacity, and low nutrient availability (Bishop and Chapin 1989).
In a chronosequence study on Mount Robson in British Columbia, Blundon et al. (1993) found that Utah sweetvetch facilitated succession in glacial moraines ranging from 46 to 184 years old. At the youngest moraines, grayleaf willow (Salix glauca) and Engelmann spruce (Picea engelmannii) were positively associated with the presence of Utah sweetvetch clumps. Utah sweetvetch presence was also related to higher rates of recruitment of later successional woody species. The study did not determine whether faciliation was through nutritional or physical factors (Blundon et al. 1993).
Utah sweetvetch growth is reduced with shading, suggesting it may be restricted to openings in late-seral communities (Spellman and Wurtz 2011). In early seral floodplain study sites along the Healy and Nenana rivers in Alaska, Utah sweetvetch biomass was significantly reduced with shading at medium, medium-high and high shade levels (62-84% obstruction of ambient solar radiation, P < 0.05). Total biomass was reduced more than 60% at medium-high shading (Spellman and Wurtz 2011).
Seed And Seedling Ecology
Utah sweetvetch is considered a good seed producer (Rumbaugh 1983). A single average-size plant growing with minimal competition in North Dakota produced 125 seeds. Seeds were harvested when production was likely at a maximum (Stevens 1957).
Seed production can be impacted by insect predation (Swoboda 2007). Butterfly and moth larvae (Lycaenidae, Gelechiidae) and adult grasshoppers consumed Utah sweetvetch flowers at several sites in Utah and Wyoming (Swoboda 2007). The hard seeds produced by Utah sweetvetch are sought out and possibly cached by rodents. Uneaten seeds carry over in the soil (Kitchen 1994).
Boukheloua (1990) found that Utah sweetvetch seedlings grew slowly. Plants averaged 0.64 g/plant after several months in a greenhouse. Jacobs and Sheley (1999) similarly reported slow seedling growth of Utah sweetvetch harvested six times at 14-day intervals (Table 2). Measurements began 14 days after seedlings were thinned to one/pot (6 in [15 cm] diameter, 39 in [100 cm] deep). Pots were filled with 2/3 Farland silt loam and 1/3 sand and initially seeded 2 mm deep with 10 seeds. Seedlings were grown in a greenhouse at 54 °F (12 °C) with a 12-hr light/dark cycle (Jacobs and Sheley 1999). Growth may be more rapid in natural settings as Northstrom (1969) reported that in the field taproots can reach greater than 3 feet (1 m) in the first growing season.
Table 2. Growth rates of Utah sweetvetch seedling roots, leaves, and shoots as determined from linear regression equations of Utah sweetvetch seedlings grown in a greenhouse (Jacobs and Sheley 1999).
| Attribute | Root length (mm/d) | Leaf area (cm²/d) | Root weight (mg/d) | Shoot weight (mg/d) |
| Growth rate | 73 | 4.1 | 2.1 | 1.6 |
| R2 | 0.5 | 0.2 | 0.7 | 0.7 |
Disturbance Ecology
References to the disturbance tolerance of Utah sweetvetch are mostly observational or anecdotal. Although many viewed Utah sweetvetch as tolerant of grazing, even heavy grazing (USDA FS 1937; Hermann 1966; Redente 1980), Wasser and Shoemaker (1982) considered it only fairly or weakly tolerant to grazing and reported reduced abundance on many grazed Utah ranges. In prairie vegetation of northern Kananaskis County, western Alberta, the abundance of Utah sweetvetch was greater on ungrazed than on grazed plots. Sites were moderately grazed by feral horses in late summer and fall for 25 years. Utah sweetvetch cover did not differ between grazed (0.84%) and ungrazed plots (1.26%), however, frequency was significantly less (P < 0.05) on grazed (2%) than ungrazed (18%) plots (Catling et al. 2015).
Utah sweetvetch tolerates fire when plants are dormant (Wasser and Shoemaker 1982). Its abundance increased strongly following stand-replacing fires or clearcutting followed by broadcast burning in Douglas-fir (Pseudotsuga menziesii) and white fir (Abies concolor) forests in western Montana. Sites were visited 4 to 80 years after wildfire and treatments (Arno et al. 1985).
Wildlife And Livestock Use
Utah sweetvetch provides excellent forage for wildlife and all classes of livestock, and it is an important pollinator plant (Hermann 1966; Stevens 2004a; Rosales 2014). Utah sweetvetch roots are eaten by meadow voles (Microtus spp.) and lemmings (Lemmini spp.). Domestic dogs that find the root caches made by rodents will eat the roots (Moerman 2003).
Plants produce green growth early in spring and the highly palatable forage remains succulent through the growing season. Some basal leaves remain green through the winter (Plummer et al. 1968). The forage contains condensed tannins, which make it a bloat-safe legume (Rumbaugh 1983). In Italy, Algeria, Spain, and other parts of the Mediterranean, a Utah sweetvetch relative, sulla (Hedysarum coronarium), is cultivated like alfalfa (Medicago spp.) (USDA FS 1937).
Utah sweetvetch is highly nutritious and eaten by bison (Bison bison), moose (Alces alces), elk (Cervus canadensis), deer (Odocoileus spp.), and pronghorn (Antilocapra americana) (Stevens 2004a; Rosales 2014). Utilization by big game is heavy in some seasons (Stevens 2004a). Utah sweetvetch was also an important and highly selected food for reintroduced muskox (Ovibos moschatus) in northeastern Alaska, even though it was sparse in the study area (Robus 1981). It was also considered a first-class forage and highly palatable to cattle, goats, and sheep on the Wasatch National Forest in Utah (Hermann 1966).
Utah sweetvetch is a high-quality food for grizzly bears (Ursus arctos) that dig and eat its roots (Doak and Loso 2003). In feeding observations in Alaska, grizzlies typically left portions of Utah sweetvetch roots sufficient for plant regrowth (Robus 1981).
In western Alberta, Pigeon et al. (2016) found that increased abundance of sweetvetch (Hedysarum spp.) was a criterion for den site selection by grizzly bears. In a 2000 to 2021 study of 10 male and 21 female grizzly bears, den sites with greater cover and presence of Utah sweetvetch were selected because of its high value as spring forage. The probability of den selection increased with increased cover of sweetvetch (Pigeon et al. 2016). This selection factor was second only to den concealment by canopy cover.
Greater sage-grouse (Centrocercus urophasianus) eat the flowers and leaves of Utah sweetvetch. The species is considered an excellent sage-grouse food source and provides moderately good cover (Hess 2011; Rosales 2014; Luna et al. 2018).
In an assessment of the pollinator friendliness of plant species, Utah sweetvetch rated high for species richness and abundance of bee visitation (Glenny et al. 2022). It rated low for uniqueness of visiting bee communities with just one bee specialist visitor. It also rated low for its spatial and temporal availability of floral resources, which were available in the fewest number of sites from mid-June to mid-July. The methods used to determine pollinator friendliness used observations of plant-bee interactions in the 2018 and 2019 growing seasons at sites covering a range of environmental conditions in the Lewis and Clark National Forest in west-central Montana (Glenny et al. 2022).
Utah sweetvetch is a host plant for a variety of moths, butterflies, and bees (James and Nunnallee 2011; Discover Life 2024; Robinson et al. 2025). It is a host plant for the following lepidopterans: Biston betularia, Coleophora accordella, Colias alexandra, C. hecla, C. philodice, Glaucopsyche lygdamus, Lycaeides melissa, and Plebejus saepiolus (James and Nunnallee 2011; Robinson et al. 2025) and a host plant for a large variety of bee species (Table 3).
Table 3. Bee species reported to use Utah sweetvetch as a host plant (Tepedino and Stackhouse 1987; Swoboda 2007; Discover Life 2024).
| Bee Taxa | Tepedino and Stackhouse 1987 | Swoboda 2007 | Discover Life 2024 |
| ANDRENIDAE | |||
| Andrena spp. | X | ||
| APIDAE | |||
| Anthophora porterage | X | ||
| Anthophora ursina | X | ||
| Apis mellifera | X | X | X |
| Bombus appositus | X | X | X |
| Bombus bifarius | X | ||
| Bombus fervidus | X | X | X |
| Bombus flavifrons | X | X | |
| Bombus griseocollis | X | X | |
| Bombus huntii | X | ||
| Bombus insularis | X | X | |
| Bombus nevadensis | X | X | X |
| Bombus occidentalis | X | ||
| Bombus rufocinctus | X | X | |
| Bombus sylvicola | X | X | |
| Eucera frater | X | X | X |
| Habropoda morrisoni | X | ||
| Nomada spp. | X | ||
| HALICTIDAE | |||
| Evylaeus spp. | X | ||
| Halictus spp. | X | ||
| Halictus rubicundus | X | ||
| MEGACHILIDAE | |||
| Callanthidium formosum | X | ||
| Hoplitus albifrons | X | X | |
| Hoplitus fulgida | X | ||
| Hoplitus hypocrita | X | ||
| Hoplitus producta | X | X | |
| Hoplitus sambuci | X | ||
| Megachile frigida | X | X | X |
| Meachile gemula | X | X | X |
| Megachile inermis | X | X | |
| Megachile melanophaea | X | X | X |
| Megachile perihirta | X | ||
| Osmia albolateralis | X | X | X |
| Osmia atrocyanea | X | X | X |
| Osmia bruneri | X | X | |
| Osmia bucephala | X | X | X |
| Osmia cockerelli | X | X | |
| Osmia cyanella | X | ||
| Osmia dolerosa | X | ||
| Osmia ednae | X | ||
| Osmia grindeliae | X | X | X |
| Osmia inermis | X | X | |
| Osmia integra | X | ||
| Osmia lignaria | X | ||
| Osmia longula | X | X | |
| Osmia nifoata | X | X | |
| Osmia paradisica | X | X | X |
| Osmia pentstemonis | X | X | |
| Osmia proxima | X | X | |
| Osmia pusilla | X | X | X |
| Osmia simillima | X | ||
| Osmia tersula | X | X | |
| Osmia tristella | X | X |
Nutritional Value
Several studies provide nutritional information for Utah sweetvetch forage and flowers (Redente 1980; Johnson et al. 1989). One year of measurements at a site in northwestern Colorado found that digestible protein of Utah sweetvetch is high during its early growth but decreases substantially when the plant is fruiting. Digestible energy of Utah sweetvetch met the minimum nutrient requirements of cattle and sheep in the vegetative growth phase (Redente 1980).
Growth and nutritive characteristics were evaluated for 11 Utah sweetvetch ecotypes grown in a common garden in Logan, Utah (Table 4, Johnson et al. 1989). Characteristics were evaluated from harvests made in mid-June and early October in 1984 on plants that were transplanted to the field in June 1983. There were significant differences (P < 0.05) in forage yields and invitro dry matter digestibility by ecotype in June but not in October. Crude protein varied among ecotypes only in October (Johnson et al. 1989).
Table 4. Range and average values for growth and nutritive characteristics of 11 Utah sweetvetch ecotypes grown in a common garden in Logan, Utah (Johnson et al. 1989).
| Harvest date | June range | June average | October range | October average |
| Forage weight (g/plant) | 41-168 | 95 | 45-115 | 83 |
| Crude protein (%) | 12.9-17.3 | 15.9 | 5.4-10.3 | 8.3 |
| IVDMD (%)¹ | 43.9-54.8 | 50.4 | 28.8-49.3 | 37.7 |
¹Invitro dry matter digestibility.
Forage production of two Utah sweetvetch cultivars (‘Basin’ and ‘Timp’) were compared at common garden sites in Utah (Table 5a; Peel et al. 2023). Nutritive value of the cultivars was also compared with alfalfa (Table 5b). Basin and Timp seedlings were grown in a greenhouse then spring outplanted in a common garden in Millville, Utah. Basin consistently produced more forage than Timp (P < 0.05), and within three years, forage production was 75% greater for Basin than Timp (Table 5b). At another common garden in Lewiston, Utah, regrowth following plant harvests in June was compared for 2 years. The average regrowth production of Basin (386 lb/ac [433 kg/ha]) was significantly greater than for Timp (227 lb/ac [255 kg/ha]) (P < 0.05).
Table 5a. Forage biomass (lb/ac) of two Utah sweetvetch cultivars grown in a common garden in Millville, Utah. Biomass of seedlings outplanted in spring 2012 was measured for three years in June (Peel et al. 2023).
| Year | 2013 | 2014 | 2015 | Average |
| Basin | 1,459a¹ | 1,376a | 1,274a | 1,369a |
| Timp | 822b | 775b | 728b | 775b |
|
Year |
2013 |
2014 | 2015 |
Average |
| ……………………………..lb/ac…………………………….. | ||||
| Basin UT sweetvetch |
1,459a1 |
1,376a | 1,274a |
1,369a |
| Timp UT sweetvetch |
822b |
775b | 728b |
775b |
1Values in the same column with different letters differ significantly (P ≤ 0.05).
Crude protein of Basin and Timp regrowth was similar significantly greater than ‘Vernal’ alfalfa (M. sativa) in the spring, but significantly lower than Vernal alfalfa in the fall (Table 5b). Neutral detergent fiber values of spring and fall regrowth were higher for Vernal alfalfa than for either Utah sweetvetch cultivar.
Table 5b. Nutritive value of spring (June) and fall (September) regrowth of Utah sweetvetch cultivars and Vernal alfalfa growing in a Lewiston, Utah, common garden (Peel et al. 2023).
| Species and cultivar | Crude protein (g/kg) | NDF* (g/kg) | ||
| June | Sept (regrowth) | June | Sept (regrowth) | |
| Basin UT sweetvetch | 294a | 146b | 247b | 300a |
| Timp UT sweetvetch | 285a | 165ab | 279a | 294a |
| Vernal alfalfa | 218b | 178a | 296a | 372b |
| Species and cultivar | Crude protein (g/kg) | NDF* (g/kg) | ||
| June | Sept (regrowth) | June | Sept (regrowth) | |
| Basin UT sweetvetch | 294a | 146b | 247b | 300a |
| Timp UT sweetvetch | 285a | 165ab | 279a | 294a |
| Vernal alfalfa | 218b | 178a | 296a | 372b |
1Neutral detergent fiber digestibility. 2Values in the same column with different letters differ significantly (P ≤ 0.05).
Several sources reported the nutritional content of Utah sweetvetch flowers and seeds. Swoboda (2007) measured 20 Utah sweetvetch flowers on plants growing in Utah. Flowers averaged 0.24 µl of nectar/flower (range: 0.08-0.46 µl), and the nectar averaged 60% sugar content (range: 45-66%). In montane and subalpine sites in the Kananaskis Valley in Alberta, Utah sweetvetch flowers averaged 0.13 mg nectar sugar/flower (Kudo and Harder 2005). Utah sweetvetch seeds average 8.8% oil and 43% protein (SER, INSR, RBGK 2024).
Ethnobotany
Utah sweetvetch roots are nutritious and eaten by Indigenous peoples (Hermann 1966; Taylor 1992; Shaw 1995). Roots were gathered in spring and eaten by Indians and early European colonizers (Craighead et al. 1963; MacKinnon et al. 2014). Indigenous peoples in the Great Basin reported that the roots taste like licorice (Taylor 1992). MacKinnon et al. (2014) reports that young Hedysarum spp. roots have a sweet licorice taste when eaten raw and taste like carrots when cooked. Roots can be boiled, baked, fried, or cooked in soups or stews. Hot drinks are made by adding a fried piece of root to hot water. Pieces of roots that were chewed and softened served as pacifiers for babies. Preserved roots were used in trading (MacKinnon et al. 2014).
Horticulture
Utah sweetvetch plants and seed are available commercially (Utah State University 2024). Its bright showy flowers, drought tolerance, pollinator attractiveness, and low maintenance requirements make it a good choice for gardens, roadside improvements, campgrounds, or other native plant landscapes in USDA Hardiness Zones 3 to 7 (Mee et al. 2003; Meyer et al. 2009; Peel et al. 2011; Rosales 2014; PFAF 2024). Utah sweetvetch can be used to add beauty and diversity to sites receiving low precipitation (Rosales 2014). It self-sows, exhibits moderate seedling vigor, grows at a moderate rate, and is pollinated by native bees and bumble bees, making it valuable for use in meadows (Mee et al. 2003), though it can also be planted in more formal perennial beds. Specimens purchased from a nursery may require acclimatization prior to outplanting (Meyer et al. 2009).
Utah sweetvetch made the “Utah’s Choice” list, a plant tagging and education program designed to promote native plants and the Intermountain Native Plant Growers Association (Meyer 2005). The committee chose 40 species from the large number of native Intermountain species worthy of landscape use. Species were chosen primarily for their outstanding beauty, ease of propagation, mention in native plant lists for the Great Basin, and proven success in a landscape setting. This program was a marketing effort to create demand for these plants while increasing and stabilizing native plant supplies generally (Meyer 2005).
Revegetation Use
Utah sweetvetch is an important wildlife and livestock forage and pollinator species with many growth characteristics that make it a good candidate for use in revegetation. It provides erosion control; tolerates grazing, drought, and other disturbances; grows in saline and alkaline soils; and persists once established (Ogle et al. 2011; Rosales 2014; EcoRestore AZ 2024; Utah State University 2024). It is capable of good seed production and high levels of germination in habitats receiving 9 to 18 in (23-46 mm) of annual precipitation (Rumbaugh 1983; Jensen et al. 1999; Ogle et al. 2011; Utah State University 2024). It is also a good soil stabilizer and capable of fixing nitrogen, which is eventually released into the soil and improves soil quality (Rosales 2014).
Developing A Seed Supply
For restoration to be successful, the right seed needs to be planted in the right place at the right time. Coordinated planning and cooperation is required among all partners to select appropriate species and seed sources; these are determined by site conditions and current or projected revegetation requirements and goals (PCA 2015) and consultation with land managers and stakeholders.
Developing a seed supply begins with seed collection from native stands. Collected seed is distributed for restoration or production of nursery stock. Increase in agricultural seed fields may be required to provide a supply of seed (PCA 2015). Production of nursery stock requires less seed than large-scale seeding operations. Regardless of the size and complexity of any revegetation effort, seed certification is essential for tracking seed origin from collection through use (UCIA 2015).
Seed Sourcing
Utah sweetvetch populations showed high levels of diversity based on the population structure and genetics of 12 wildland collections, four accessions from the National Plant Germplasm System (Pullman, WA), and ‘Timp’ a release developed from Intermountain West accessions (Bushman et al. 2007). These populations and accessions represented four main genetic clades: Alaska, Idaho, western Utah, and eastern Utah and neighboring Colorado. The two Utah clades were separated by the Wasatch Mountains. Accessions had high diversity with 70% of the genetic variation occurring within accessions. Northeastern Utah accessions had the highest levels of genetic diversity, but overall, differences in diversity were marginal and sometimes insignificant. High levels of diversity are common in outcrossing plants. This information was shared with those developing germplasm to inform creation of broad-based improved germplasm (Bushman et al. 2007).
Because empirical seed zones are not currently available for Utah sweetvetch, generalized provisional seed zones developed by Bower et al. (2014) may be used to select and deploy seed sources. These provisional seed zones identify areas of climatic similarity with comparable winter minimum temperature and aridity (annual heat to moisture index). In Figure 8, Omernik Level III Ecoregions (Omernik 1987) overlay the provisional seed zones to identify climatically similar but ecologically different areas. For site-specific disturbance regimes and restoration objectives, seed collection locations within a seed zone and ecoregion may be further limited by elevation, soil type, or other factors.
The Western Wildland Environmental Threat Assessment Center’s (USDA FS WWETAC 2024) Threat and Resource Mapping (TRM) Seed Zone application provides links to interactive mapping features useful for seed collection and deployment planning. The Climate Smart Restoration Tool (Richardson et al. 2020) can also guide revegetation planning, seed collection, and seed deployment, particularly when addressing climate change considerations.
Occurrence Map
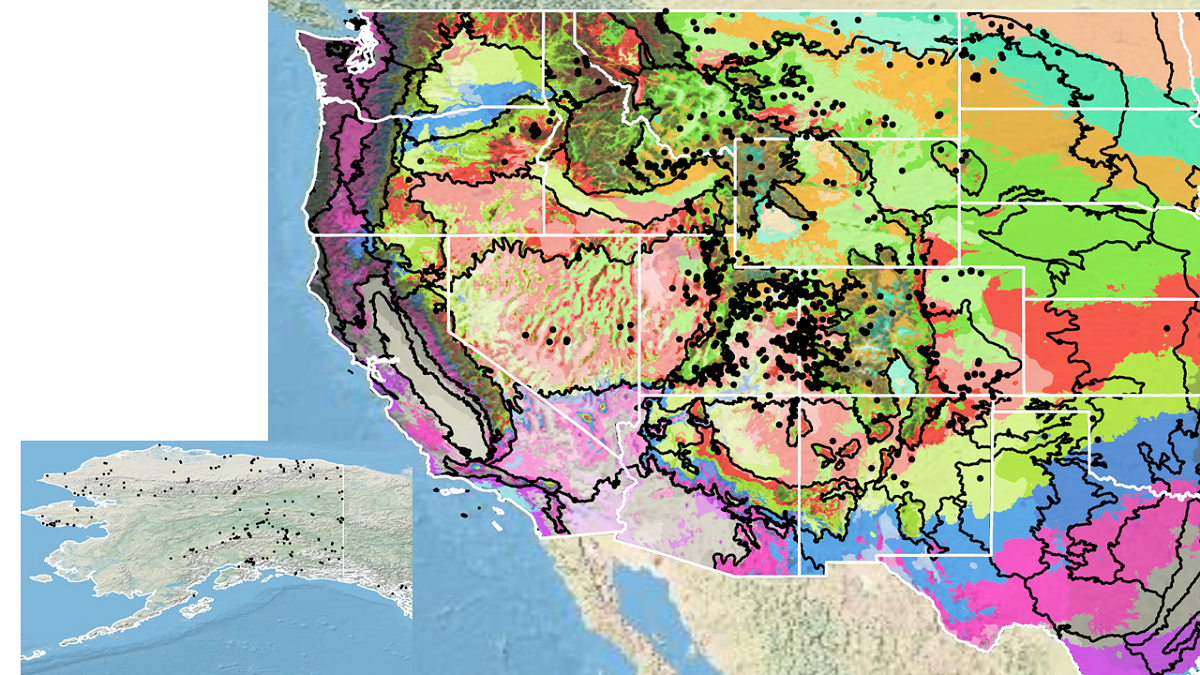
Figure 8. Distribution of Utah sweetvetch (black circles) based on geo-referenced herbarium specimens and observational data from 1869-2023 (CPNWH 2017; SEINet 2017; USDI USGS 2023). Generalized provisional seed zones (colored regions) (Bower et al. 2014) are overlain by Omernik Level III Ecoregions (black outlines) (Omernik 1987; USDI EPA 2018). Interactive maps, legends, and a mobile app are available (USDA FS WWETAC 2017; https://www.fs.usda.gov/wwetac/Seedzonemapping.php). Map prepared by Sarah Barga and Benton Spendlove USFS RMRS.
Releases
‘Timp’ and ‘Basin’ are the currently available (2025) cultivars of Utah sweetvetch (Rosales 2006). Timp is a seed-propagated cultivar from genetic material originating from a site at the base of Utah’s Wasatch Mountains and a single plant selection made by Robin Cuany (Rosales 2006). Timp was selected based on its seedling vigor, site adapatability, persistence, seed production, nitrogen-fixation, and stability. It was released in 1994 by the Upper Colorado Environmental Plant Center, Utah Division of Wildlife Resources, Colorado State University Agricultural Experiment Station, Utah State University Agricultural Experiment Station, and the USDA Natural Resources Conservation Service (Rosales 2014). Timp is adapted to well-drained loamy soils where annual precipitation is 12 to 18 in (305-457 mm) in the Intermountain region of Utah, Colorado, and Wyoming (Rosales 2006, 2014).
The Basin cultivar was released in 2021 and represents a 24-plant synthetic cultivar from three cycles of genetic selection for improved seed and forage production (Peel et al. 2023). Basin was developed to replace Timp, which has poor seed production. Parent contributions came from the earlier Timp germplasm release and eight populations in western and eastern Utah; Jefferson County, Colorado; Custer and Jefferson Counties in Idaho; and Alaska. Basin is recommended for much of the Intermountain West (Great Basin [MLRA 25,28A,28B], Snake River Plain [MLRA 10,11,12], Eastern Idaho Plateau [MLRA13], Cool Central Desert Plateaus [MLRA 34A], and Rocky Mountains [MLRA 43B]) (Peel et al. 2023).
Wildland Seed Collection
Utah sweetvetch produces seed in pods or loments (Fig. 9), which appear 2 to 3 weeks after flowering (Redente 1980). Loments remain on the plant for just 2 to 4 weeks once seeds are mature. They are easily hand collected (Northstrom 1969; Redente 1980; Winslow 2002).

Figure 9. Utah sweetvetch plant growing in Alaska with mature seed pods or loments. Photo: DOI, BLM AK930 SOS.
Wildland Seed Certification
Verification of species and tracking of geographic source is necessary whether wildland seed is collected for immediate project use or as stock seed for cultivated increase. This official Source Identification process can be accomplished by following procedures established by the Association of Official Seed Certifying Agencies (AOSCA) Pre-Variety Germplasm Program (UCIA 2015; Young et al. 2020). Wildland seed collectors should become acquainted with state certification agency procedures, regulations, and deadlines in the states where they collect.
If wildland-collected seed is to be sold for direct use in ecological restoration projects, collectors must apply for Source-Identified certification prior to making collections. Pre-collection applications, site inspections, and species and seed amount verification are handled by the AOSCA member state agency where seed collections will be made (see listings at AOSCA.org).
If wildland seed collected by a grower or private collector is to be used as stock seed for planting cultivated seed fields or for nursery propagation (See Agricultural Seed Certification section), detailed information regarding the collection site and collecting procedures must be provided when applying for certification. Photos and herbarium specimens may be required. Germplasm accessions acquired within established protocols of recognized public agencies, however, are normally eligible to enter the certification process as stock seed without routine certification agency site inspections. For contract grow-outs, however, this collection site information must be provided to the grower to enable certification.
Collection Timing
Seed pods (loments) appear 2 to 3 weeks after anthesis (Northstrom 1969; Redente 1980). Loments are tan when mature. They are retained on the plant for up to 4 weeks after which time they are dropped and begin to dehisce (Redente 1980; Winslow 2002). Seeds are reddish brown at maturity (Fig. 10) (Lapp et al 2001). Seed harvested at physiological maturity requires drying before storage. Delaying harvests until seed pods are dry and brittle, though, results in some seed loss to shattering (Peel et al. 2011).

Figure 10. Clean Utah sweetvetch seeds under magnification. Photo: U.S. Department of Agriculture, Forest Service, Bend Seed Extractory.
The earliest seed collections made by the Department of the Interior, Bureau of Land Management’s Seeds of Success (SOS) collection crews were made on June 17, 2014, in Uintah County, Utah, at an elevation of 5,594 ft (1,705 m) (USDI BLM SOS 2025). The latest collection was made on August 30, 2005, in Garfield County, Utah, at an elevation of 8,059 ft (2,456 m). The SOS teams made 25 collections from 2002 to 2016 over an elevation range of 711 to 8,059 ft (217-2,456 m) in Utah (10), Colorado (10), Alaska (3), Idaho (1), and North Dakota (1). Most collections were made in July (16) and the rest were made in June (3) or August (6). At four collection sites there were at least two collections made between 5 days and 2 weeks after the first (USDI BLM SOS 2025). SOS records generally agree with Jorgensen and Stevens (2004) who reported that seed typically matures in early July to early August.
Collection Methods
Utah sweetvetch seed pods can be collected by hand-stripping, knocking seed into containers, vacuuming, or by combining (Plummer et al. 1968; Jorgensen and Stevens 2004). Methods will depend on the size and locations of the stand and equipment availability (Wasser and Shoemaker 1982). Eddleman (1977) reported short retention times for Utah sweetvetch loments during harvests made in 1975 and 1976, and because of this, vacuuming pods from the ground was better and faster than hand stripping (Eddleman 1977).
Several collection guidelines and methods should be followed to maximize the genetic diversity of wildland collections: 1) collect seed from a minimum of 50 randomly selected plants; 2) collect from widely separated individuals throughout a population without favoring the most robust or avoiding small stature plants; and 3) collect from all microsites including habitat edges (Basey et al. 2015). General collecting recommendations and guidelines are provided in online manuals (e.g., ENSCONET 2009; USDI BLM SOS 2023).
It is critical that wildland seed collection does not impact the sustainability of native plant populations. Collectors should take no more than 20% of the viable seed available at the time of harvest (USDI BLM SOS 2023). Additionally, care must be taken to avoid the inadvertent collection of weedy species, particularly those that produce seeds similar in shape and size to those of Utah sweetvetch.
Collection Rates
Based on seed harvesting done by the Bridger Plant Materials Center (PMC), an average of 2.7 oz (76 g) of clean seed can be collected per person-hour. Collection rates varied by year, stand density, and collector experience (Winslow 2002).
Post-Collection Management
Seed should be stored in breathable containers in a well-ventilated drying shed prior to cleaning (Lapp et al. 2001) or spread out on a tarp in a dry, sheltered environment and turned daily until no moisture or warmth is detected (3-5 days) (Winslow 2002).
Seed Cleaning
Removing seeds from tightly constricted Utah sweetvetch seed pods can be challenging (Fig. 11). Mechanical abrasion and heat from equipment can damage the seed (Eddleman 1977; Stevens et al. 1996). Most procedures detailed in the literature suggest cleaning dry seed using a debearder or hammermill to remove the seeds from the loment segments then passing the seed through an air screen separator to remove debris (Stevens et al. 1996; Jorgensen and Stevens 2004). Staff at Glacier National Park reported 100% purity and 60% germination after cleaning seed with a hammermill followed by screening (Lapp et al. 2001).

Figure 11. Utah sweetvetch seeds (dark) and individual loment segments that contain a single seed (light) or may be empty. Seed collected from plants in Wyoming. Photo: DOI, BLM WY050 SOS.
Staff at the Bridger PMC processed dry seed by threshing it through a hammermill with a 12/64 in round-hole screen (Winslow 2002). It was then air-screened using a clipper M2B or eclipse cleaner using a 17/64 in round-hole screen and moderate air speed. Because of its large seed size, lack of seed debris, and fair seed flow, Bridger PMC considers Utah sweetvetch moderately easy to clean. Large seed lots are efficiently processed using mechanized equipment, and small seed lots require more hand labor (Winslow 2002). Wasser and Shoemaker (1982) cleaned seed using an air blaster equipped with sieves, followed by screening or processing through a fanning mill.
Estimates of seed quality made during the seed cleaning process can be helpful to determine when recleaning is needed to improve purity (Tilley and Ogle 2011). Historic bushel weights can be useful in estimating seed quality. Seed lots that fail to meet expected bushel weights can be recleaned with higher air flow rates to remove lighter, unfilled seed, and increase the percentage of viable seed. Bushel weight comparisons can be made by weighing the amount of seed that fills a standard US measuring cup (237 ml). The bushel weight calculation is straightforward using English units, since a bushel contains approximately 149 cups. So, lb/bushel = (ounces per cup * 149)/16. For comparison, the lbs/bushel weight for Timp Utah sweetvetch is 50 (Tilley and Ogle 2011).
Seed Storage
Utah sweetvetch seed can be stored in or out of the pod. Treatments are necessary to control insects (Stevens and Monsen 2004). Managing bruchid beetle larvae feed on legume seeds is essential to avoid seed damage during storage (Peel et al. 2011). Without treatment, more than 50% of a small seed lot was lost in 3 months of storage, even though beetles were not observed after plants were treated with insecticide after flowering (Peel et al. 2011).
At Glacier National Park, Montana, Utah sweetvetch seed longevity was retained for up to 10 years when seed was kept in sealed containers at 34 to 37 °F (1-3 °C) (Lapp et al. 2001). Other researchers reported longevity of up to 6 years for seed stored without special care and exposed to widely fluctuating temperatures (Plummer et al. 1968; Stevens et al. 1996; Jorgensen and Stevens 2004). Redente (1980) reported 94% viablity for seed stored for 1 to 3 years and 65% viability after 18 years.
Stevens and Jorgensen (1994) tested germination of Utah sweetvetch periodically for up to 25 years of storage in an open warehouse in Ephraim, Utah (Table 6, Stevens and Jorgensen 1994). Germination significantly decreased (by about 50%) between years 5 and 7. Seed was cleaned to at least 85% purity and stored in cotton bags in a filing cabinet housed in a warehouse where temperatures ranged from -22 °F (-30 °C) to 101 °F (38 °C). Germination was tested for 100 seeds placed between damp papers that were wrapped in plastic and kept in a refrigerator at 34 to 38 °F (1-3 °C) (Stevens and Jorgensen 1994).
Table 6. Germination percentages for Utah sweetvetch seed stored 2 to 25 years in an open warehouse in Ephraim, Utah, where temperatures fluctuated from -22 to 101 °F (-30 to 38 °C) (Stevens and Jorgensen 1994).
| Years of storage | 2 | 3 | 4 | 5 | 7 | 10 | 15 | 20 | 25 |
| Germination (%) | 59 | 67 | 58 | 55 | 25 | 40 | 16 | 11 | 21 |
Seed Testing
The quantity of seed required for testing is 0.7 oz (20 g) for purity and germination testing and 7 oz (200 g) for noxious weed count (Belcher 1985).
Viability Testing
There are Association of Official Seed Analysts (AOSA 2010) procedures and guidance for testing Utah sweetvetch viability. To prepare the seed for testing, allow the seed to imbibe water overnight by placing it between moist media at temperatures of 68 to 77 °F (20-25 °C). To test the viability of hard seed, clip or nick the seed coats of seeds that did not imbibe moisture. Put imbibed and/or nicked seeds into a 1% tetrazolium (TZ) solution for 2 to 72 hours at 86 to 95 °F (30-35 °C). Seed should be cleared with glycerol or lactic acid for about 1 hour at 95 °F (35 °C) before evaluating staining (AOSA 2010).
Seed is viable if:
- the entire embryo is evenly stained, turgid, and unfractured,
- unstained areas of the radicle or hypocotyl do not extend into conducting tissue,
- the radicle stains slightly darker,
- there are small, shallow, unstained or intensely stained areas on the periphery of the hypocotyl and cotyledons, or
- if half or more of the cotyledons attached to the embryo axis are evenly stained (AOSA 2010).
Seed is not viable if:
- the embryo is completely unstained,
- the embryo is flaccid,
- the radicle is unstained, deteriorated, or fractured above the tip of central conducting tissue,
- there are unstained, watery, or darkly stained areas extending into the inner surfaces of the cotyledons,
- less than half of the cotyledons remain functional and attached to the embryo axis, or
- the embryo axis is fractured or necrotic (AOSA 2010).
Germination Testing
There is AOSA (2023) guidance for testing germination. Seed should be kept moist and incubated at alternating 59 to 77 °F (15-25 °C) or constant 68 °F (20 °C). The first germination count is made on day 14, and the last germination count is made on day 28. A portion of the seeds are normally hard and impermeable to water. The percentage of hard seed should be reported with the percentage that germinated (AOSA 2023).
Germination Biology
Maximum germination requires 1 to 2 months of afterripening and up to 4 weeks of cool, moist stratification (Stevens et al. 1996; Jorgensen and Stevens 2004; Rosales 2014). Utah sweetvetch produces a high proportion of hard seeds (Wasser 1982). Once hard seed coats are cracked, germination time is reduced by 50 to 70% (Northstrom 1969). Laboratory germination of 98 to 100% was achieved by partial removal or chipping of the seed coat, placing the seed on 1% agar, and exposing it to an 8 h/16 hr light-dark cycle at temperatures of 50 to 59 °F (10-15 °C). Germination counts were made at 7 and 21 days (SER, INSR, RBGK 2024).
Several scarification procedures have been tested for Utah sweetvetch. Boukheloua (1990) reported germination of about 95% for seed treated with concentrated sulfuric acid for 10 minutes or rubbed between sandpaper until their shine was dulled. The seed was collected near Fort Collins, Colorado, in 1986 and 1987 and germinated in the dark at 68 °F (20 °C) (Boukheloua 1990). Dreesen and Harrington (1997) reported that although there were no significant differences by treatment, germination was lowest (32-39%) for mechanical and highest (52%) for hot water scarification. The untreated seed germinated at 37%. Mechanical treatments involved processing with a Forsberg scarifier lined with 100-grit sandpaper for 3 to 75 seconds. Seed was soaked 4 hours in 194 °F (90 °C) water in the hot water treatment. The study provided no details about seed source or storage conditions or duration prior to treatments (Dreesen and Harrington 1997).
Although Utah sweetvetch seed can be stored and sold in the pods (Stevens et al. 1996), germination is improved by pod removal. Jorgensen and Wilson (2004) evaluated 20 accessions of shelled seeds and 10 accessions of unshelled seeds. Seed was incubated in the dark at 34 to 38 °F (1.1-3.3 °C). Germination of shelled Utah sweetvetch was 63% after 200 days, and 34% for unshelled seeds after 210 days.
When the effects of temperature, light, and moisture on scarified Utah sweetvetch seed were evaluated, germination was better at moderate than at extreme temperatures, faster at higher than at lower temperatures, better in dark than light, and significantly reduced by moisture stress of -10 bars osmotic pressure (Redente 1980, 1982). The study tested three seed sources: Salt Lake City, UT, 1976; Orem, UT, 1978; Musselshell, MT, 1960. All seed was mechanically scarified using a small metal drum with sandpaper and 35 psi air pressure before testing the effects of temperature, light, and moisture on germination. Mid-range temperatures of 50, 59, 68, and 77 °F (10, 15, 20, 25 °C) resulted in significantly better germination than the extremes of 41 °F (5 °C) and 86 °F (30 °C) (P = 0.01). Germination was better for younger seed collected in Utah that had higher seed viability (94%) than older seed collected in Montana (65% viability). Averaged over all temperatures, germination was significantly better in complete dark than with 8 hours of light (P = 0.01). Germination rates were slowest at the lowest temperatures (8 days to 50% at 5 °C and 7 days at 10 C). Germination was faster at higher temperatures and decreased significantly at moisture stress of -10 bars osmotic pressure or more. Germination was 70% at -1/3 bars, 50% at -5 bars, 42% at -7.5 bars then reduced markedly, 22% at -10 bars and 4.2% at -12.5 and -15 bars of osmotic pressure (Redente 1980,1982).
In another study of the effects of seed age, temperature, light, and stratification on germination of Utah sweetvetch, higher temperatures, stratification, and light improved germination (Table 7, (Eddleman 1977). Germination was best (82%) for newer seed exposed to light and higher temperatures (68 °F [20 °C]) or for 1-year-old stratified seed that was exposed to light and higher temperatures (alternating 68-86 °F [20-30 °C]).
Table 7. Germination percentages and rates for different aged Utah sweetvetch seed exposed to various temperature and light conditions (Eddleman 1977).
| Seed age (months)¹ | Pretreatment | Temp (°F) | Light (hrs) | Germination (%) | Days to 50% germination |
| 2 | None | 50 | Dark, no light | 18 | 25 |
| 2 | None | 68 | Dark, no light | 34 | 9 |
| 4 | None | 68 | 14 | 82 | 5 |
| 12 | None | 50 | Dark, no light | 56 | 11 |
| 12 | Stratification: 3 mo at 39 °F | Alternating 86 °F light, 68 °F dark | 14 | 82 | 1 |
¹All seed stored dry at 68 °F.
Wildland Seed Yield And Quality
Post-cleaning seed yield and quality of seed lots collected in the Intermountain region are provided in Table 8 (USDA FS BSE 2021). The results indicate that Utah sweetvetch seed can generally be cleaned to high levels of purity and seed fill and that viability of fresh seed is variable. Values reported in table 8 are slightly below the 69,533 seeds/lb (153,291 seeds/kg) of total clean seed (not pure live seed [PLS]), reported by Wiese et al. (2012), which was based on reported seed weights in publications, commercial seed catalogs, and seed weights measured at Glacier National Park, Montana.
Table 8. Seed yield and quality of Utah sweetvetch seed lots collected in the Intermountain region, cleaned by the Bend Seed Extractory, and tested by the Oregon State Seed Laboratory or the USDA Forest Service National Seed Laboratory (USDA FS BSE 2021).
| Seed lot characteristic | Mean | Range | Samples (no.) |
| Bulk weight (lbs) | 3.56 | 0.44-25 | 10 |
| Clean weight (lbs) | 1.32 | 0.045-10 | 10 |
| Clean-out ratio | 0.29 | 0.058-0.60 | 10 |
| Purity (%) | 98.5 | 97-99 | 10 |
| Fill (%)¹ | 86.3 | 78-99 | 10 |
| Viability (%)² | 77.9 | 24-98 | 8 |
| Seeds/lb | 47,810 | 34,552-54,676 | 10 |
| Pure live seeds/lb | 35,852 | 12,042-53,047 | 8 |
¹ 100 seed X-ray test
² Tetrazolium chloride test
Marketing Standards
Acceptable seed purity, viability, and germination specifications vary with revegetation plans. Purity needs are highest for precision seeding equipment used in nurseries, while some rangeland seeding equipment handles less clean seed quite well.
Utah sweetvetch seed is most often marketed with the pods removed from the loment segments, although it is sometimes sold with the segments intact (Stevens et al. 1996). In the reviewed literature, accepted levels of seed purity were 90%, viability ranged from 60 to 95%, and germination ranged from 60 to 90% (Wasser 1982; Stevens and Meyer 1990; Stevens et al. 1996; Jorgensen and Stevens 2004; Walker and Shaw 2005).
Agricultural Seed Production
Utah sweetvetch seed has been produced successfully in cultivation (Stevens and Monsen 2004). Preliminary farm work on the species revealed that improper timing of irrigation and harvests, lack of pollinators, and bruchid seed beetles (Bruchidae) can limit Utah sweetvetch seed production. All factors can be controlled but proper monitoring and timing are critical (Peel et al. 2011).
Agricultural Seed Certification
In order to minimize genetic changes in specific accessions of native species when increased in cultivated fields, it is essential to track the geographic source and prevent inadvertent hybridization or selection pressure. This is accomplished by following third party seed certification protocols for Pre-Variety Germplasm (PVG) as established by the Association of Official Seed Certification Agencies (AOSCA). AOSCA members in the U.S., Canada, and other countries administer PVG requirements and standards that track the source and generation of planting stock. Field and cleaning facility inspections then monitor stand establishment, proper isolation distances, control of prohibited weeds, seed harvesting, cleaning, sampling, testing, and labeling for commercial sales (UCIA 2015; Young et al. 2020).
Seed growers apply for certification of their production fields prior to planting and plant only certified stock seed of an allowed generation (usually less than four). The systematic and sequential tracking through the certification process requires preplanning, knowing state regulations and deadlines, and is most smoothly navigated by working closely with state certification agency personnel. See the Wildland Seed Certification section for more information on stock seed sourcing.
Site Preparation
Utah sweetvetch establishes best in a firm, weed-free seedbed having good soil moisture to 4 in (10 cm) deep (Winslow 2002).
Seed Pretreatments
Inoculation of seed with appropriate Rhizobia spp. is recommended (Jorgensen and Stevens 2004).
Weed Management
Minimizing weed competition should benefit Utah sweetvetch growth and seed production. Hand weeding is recommended until plants are established. Weed control after that can be through mechanical (light cultivation) or chemical (preemergent herbicides or spot spraying) treatments (Stevens et al. 1996; Rosales 2014).
While there are no herbicides recommended for use with Utah sweetvetch seed production, preliminary greenhouse and field studies evaluated the effects of glyphosate on an experimental Utah sweetvetch cultivar (USV09) and tall fescue (Schedonorus arundinaceus), which is sensitive to glyphosate. Glyphosate was applied to 4-week-old seedlings in the greenhouse and to mature field growing plants in the spring and fall. Application rates tested were 210, 420, 841, 1681, 3363, 6725, 13,450 g ae/ha, where 841 g ae/ha is the common rate applied commercially. In the greenhouse, visible injury to Utah sweetvetch seedlings exceeded 50% but was significantly less than injury to tall fescue. Following spring treatments in the field, stunted growth of Utah sweetvetch was minimal at the three lowest application rates but reached 20% at the 1681 g ae/ha rate, whereas tall fescue was stunted more than 30% at the lowest rate. Following fall glyphosate treatments, stunted growth of Utah sweetvetch was 10% or less at all but the two highest treatment levels and never exceeded 40% (Peel et al. 2015).
Seeding
Although seed that is scarified prior to planting can be seeded in spring (Rosales 2006), late fall planting provides for natural stratification and scarification (Stevens et al. 1996; Winslow 2002; Ogle et al. 2003; Rosales 2006). Utah sweetvetch should be inoculated with a Rhizobium strain for nitrogen fixation just prior to seeding, In addition, plant growth was improved in soils supporting Glomus fasciculatus, a mycorrhizal symbiont (Wasser 1982).
Recommendations for seeding Utah sweetvetch range from 10-30 PLS/ft (32-98/m) of row with row spacings of 3 to 5 ft (1-1.5 m) (Stevens et al. 1996; Winslow 2002). It can be seeded using a single row, push-type belt seeder (Stevens et al. 1996; Winslow 2002). It can also be seeded at a rate of 3 to 4 PLS in hills spaced 2 to 4 ft (0.6-1.2 m) apart (Stevens et al. 1996).
Seeding depths of 0.13 to 0.75 in (0.3-1.9 cm) are recommended (Stevens et al. 1996; Jensen et al. 1999; Winslow 2002; Ogle et al. 2003; Rosales 2006). Wasser (1982) recommended shallower (0.25 in [0.6 cm]) seeding depths for fine-textured, moist soils; slightly deeper (0.5 in [1.3 cm]) seeding in loam soils, and deepest seeding (up to 0.75 in [1.9 cm]) in dry, coarse soils. Emergence at seeding depths of 0.2 to 2 in (0.5-5 cm) was evaluated for three Utah sweetvetch seed sources planted into field-collected loam soils and grown in a greenhouse. Emergence was similar at 0.2 to 0.8 in (0.5-2 cm) seeding depths and decreased at depths of 1 in (2.5 cm) or more (Redente 1980).
Establishment And Growth
Germination and initial establishment of Utah sweetvetch can be erratic (Stevens et al. 1996). Based on monitoring of seed production plots at Ephraim and Millville, Utah, plants required 723 growing degree days (gdd) for flowering and 1,547 gdd to produce mature seed (Peel et al. 2011). The simple heat-unit model accumulated gdds at air temperatures greater than 40 °F (4 °C). Because the model was developed in the Intermountain Region, these numbers may have limited application outside this area (Peel et al. 2011).
Fertilizer is not recommended for the establishment year because it stimulates weed growth and competition (Winslow 2002). Nitrogen additions depend on field conditions (Stevens et al. 1996; Rosales 2014). Some growers recommend up to 30 lbs/ac (33 kg/ha) of available phosphate be applied every other year once plants are established. While stand density rarely increases, some accessions are rhizomatous and require between-plant cultivation (Stevens et al. 1996).
Plants flower over an extended period from early May to mid-July (Stevens et al. 1996). The loments, which drop when dry, mature unevenly on the plants. Mature seed is often available from early July to early August.
Common garden experiments compared forage yield, nitrogen fixation, and seed production of Utah sweetvetch from eleven populations from southwestern Montana to eastern Colorado sites receiving 7 to 22 in (178-559 mm) of annual precipitation (Table 9, Ford et al. 1989; Johnson et al. 1989). Seedlings were grown in a greenhouse and then transplanted to the Utah State University Evans Experimental Farm near Logan, Utah, to evaluate candidates for breeding and commercial seed production. Transplants produced most forage in spring with yield varying by 400% among populations. Some plants flowered in the establishment year but no viable seed was produced until the second growing season. An ecotype from the Ballard Mine site in Idaho which received intermediate levels of annual precipitation produced the most seed. An ecotype from Hobble Creek, Utah, the most arid region, produced the least (Table 9). Levels of nitrogen fixation varied more than tenfold among accessions. In an analysis of taxonomic relationships of these accessions, taxonomic separations were not always related to characteristics of the collection site (Ford et al. 1989; Johnson et al. 1989).
Table 9. Forage yield, nitrogen fixation, seed yield, and seed weights of 11 Utah sweetvetch accessions growing for two seasons in a common garden near Logan, Utah (Ford et al. 1989; Johnson et al. 1989).
| Origin | Elevation (ft) | Soil texture | Mean annual ppt (in) | Yield (g/plant) | N-fixation (µ moles/h/plant) | Seed yield (g/plant) | Seed weight (mg/25 seeds) |
| Benmore, UT | 5705 | Clay loam | 13 | 54d¹ | 38bc | 4.3bc | 226de |
| Arco, ID | 5003 | Silty loam | 11 | 97bcd | 22c | 3.6bc | 211ef |
| Duchesne, UT | 5003 | Loam | 13 | 117bc | 90a | 3.2bc | 213ef |
| Ballard Mine, ID² | 6804 | Mine dump | 19 | 126ab | 81ab | 2.8bc | 271ab |
| Orem, UT | 5203 | Cobbly loam | 15 | 168a | 97a | 9.0a | 278a |
| Salt Lake City, UT | 5203 | Upland stony | 18 | 120ab | 38bc | 5.8ab | 260abc |
| Hobble Creek, UT² | 5105 | Stony loam | 17 | 122ab | 30c | 3.0bc | 272ab |
| Moab, UT | 4505 | Sandy | 7 | 64cd | 39bc | 0.1c | 255bc |
| Garden City, UT | 6004 | Mtn loam | 22 | 41d | 16c | 3.3bc | 239cd |
| Musselshell Co, MT | 5203 | Loam | 21 | 69cd | 10c | 0.2c | 149g |
| Rio Blanco Co, CO | 5705 | Loam | 13 | 64cd | 31c | 1.6bc | 199f |
¹Values within a column followed by the same letter(s) do not differ (P > 0.05).
²These ecotypes were collected from planting sites where seed of the ecotype from Orem, UT, was planted and established in 1963 at Hobble Creek and 1965 at the Ballard Mine.
Irrigation
Irrigation has been helpful during the establishment phase (Stevens et al. 1996; Rosales 2014). Providing irrigation when soil moisture is lacking at the late-bud stage and after flowering can increase seed production 3 to 8 times (Peel et al. 2011). When moisture is limited, up to 90% of seed pods may be aborted (Peel et al. 2011). Excessive moisture, though, can increase the incidence of root disease and may result in growth of fungi that produce orange or black rust spots on the foliage (Stevens et al. 1996; Peel et al. 2011). Sprinkler irrigation should be avoided when plants are flowering to promote pollination (Stevens et al. 1996). Winslow (2002) recommends providing supplemental moisture if conditions are dry following seed harvest but before freezing.
Pollinator Management
Bees are required for pollination of Utah sweetvetch flowers, and managed bee pollinators can increase seed production (Stevens et al. 1996; Peel et al. 2011). Mason bees and leafcutter bees are effective pollinators of Utah sweetvetch and thus potential candidates for use (Tepedino and Stackhouse 1987; Swoboda 2007; Peel et al. 2011). Utah sweetvetch seed production plots in Utah (Ephraim and Millville sites) showed increased seed pod formation when leafcutter bees (M. rotundata) were provided during flowering (see Table 9) (Peel et al. 2011). This bee species was chosen primarily because of its availability. Peel et al. (2011) suggested that other bees or honey bees (Apis mellifiera), may also be used to increase seed production.
Managed bee pollinators are necessary to farm Utah sweetvetch for seed in locations where local wild bee pollinators are lacking (Swoboda and Cane 2012). Bee stocking should allow for new or day-old flowers to be visited at least once, but multiple flower visits are needed to maximize seed yields. Growers should choose a bee pollinator whose foraging behavior promotes outcrossing because xenogamy is associated with increased seed viability in Utah sweetvetch (Swoboda and Cane 2012).
Swoboda (2007) found that the small-bodied bees, Bruner’s mason bee (O. bruneri) and San Rafael mason bee (O. sanrafaelae) were effective pollinators. These bees were able to nest and produce progeny with Utah sweetvetch as their only pollen and nectar source making them candidates for use in Utah sweetvetch seed production. For stocking calculations, Swoboda (2007) reported that a 2-acre (0.8 ha) stand of Timp growing near Worland, Wyoming, yielded 5,000 flowers/plant over the growing season and about 4,000,000 flowers/ac (4,483,382/ha) on an average day at mid-bloom.
If seed production fields are small and adjacent to natural habitats, Utah sweetvetch may attract enough native bees and not need managed pollinators (Tepedino and Stackhouse 1987). A three-acre (1.2 ha) field surrounded by rangeland in western Wyoming attracted an abundance of leafcutter and honey bees and produced abundant seed. But for larger fields or fields in areas where bees are scarce, Tepedino and Stackhouse (1987) developed some guidance for choosing a commercial pollinator. A solitary bee species is a potential candidate because of the small-scale nature of farming and relatively short bloom period. Thus, a candidate pollinator having one generation per year and short periods of flight is ideal. They recommended small-handed leafcutter (Megachile gemula), white-sided mason (Osmia albolateralis), Bruner’s mason, and bufflehead mason (O. bucephala) bees as worthy of testing as managed Utah sweetvetch pollinators. These bee species require minimal care and spend 10 to 11 months of the year in an immature state in their nests (Tepedino and Stackhouse 1987).
Pest Management
Utah sweetvetch has many natural predators and some fungal vulnerabilities that can impact establishment, survival, and seed production. These include a variety of small mammals, grasshoppers, crickets, seed-feeding larvae, root rot, and rusts (Wasser 1982; Stevens et al. 1996; Rosales 2014).
Several insects are potential Utah sweetvetch pests. Seed beetles (Acanthoscelides fraterculus) use Utah sweetvetch as a host plant and are a source of potential seed loss (Johnson 1990). Bruchid weevil larvae feed on developing seeds and may limit viable seed production if not controlled (Stevens et al. 1996; Peel et al. 2011). Without treatment, bruchid beetle infestations resulted in seed losses of more than 50% in a small lot stored 3 months. When plants were treated with insecticide after flowering, no beetles were observed (Peel et al. 2011). Based on sampling in small seed increase farms in southern Idaho and southern Colorado, Lygus bugs (Lygus spp.), seed beetles (Acanthoscelides spp.), Karner blue butterflies (Lycaeides melissa), and moths (Filatima xanthuris) were noted in production plots (Hammon 2007). The moths webbed and fed on flowering racemes and upper foliage of Utah sweetvetch. Infestations were persistent and affected racemes rarely produced seed. At one site, 25% of racemes were infested for two sequential years (Cane 2005).
Utah sweetvetch is also a host species for plant bugs (Lopidea bonanza, L. bullata, L. dakota, L. nigridia, L. picta, L. scutate) and fungi (Fusarium solani, Uromyces borealis, U. hedysari-obscuri, U. hedysari-paniculati) (Peck 1881; Uppala et al. 2013; Discover Life 2024). Fusarium solani infected a seed field in central Oregon where 15% of Utah sweetvetch plants exhibited chlorosis, defoliation, stunting, wilting, and/or death (Uppala et al. 2013).
Seed Harvesting
Utah sweetvetch can be harvested mechanically using a reel-type harvester or combine (Plummer et al. 1968; Stevens et al. 1996; Jorgensen and Stevens 2004). Smaller stands can be hand harvested by stripping or beating plants over a hopper or tarp (Stevens et al. 1996). Harvesting seed at its physiological maturity means collections will require careful drying before storing. Postponing harvests until seed pods are dry and brittle results in some seed loss to shattering (Peel et al. 2011).
At a seed production farm near Woodruff, Utah, plants were swathed and windrowed with a draper-type swather. Timing was important. Swathing too early resulted in harvests of shriveled seed and low viability. Swathing too late meant seed loss because fruits had shattered. Plants were cut to a height of 4.7 in (12 cm) and allowed to dry in windrows for 3 to 5 days before gathering. Using proper sieve settings, screens, and air levels, windrows were picked up and threshed with a commercial combine with minimal loss (Ford 1988).
Seed Yields And Stand Life
Utah sweetvetch produces some seed in its second year (10-35% of full production), and reaches full production in year 3. Seed yields can range from 75 to 250 lbs/ac (84-280 kg/ha) at 90% purity, and stands can be harvested for 8 years or more (Stevens et al. 1996; Rosales 2014). Optimum seed yields occur every other year (Rosales 2014).
Seed yields exceeded 250 lbs/ac (280 kg/ha) for both Basin and Timp Utah sweetvetch cultivars grown for seed production in Millville, Utah (Table 10; Peel et al. 2023). Production fields were established from greenhouse-grown seedlings transplanted in spring 2012. Nesting boards with leafcutter bee larvae were placed nearby to increase pollinator visitation. Seed was harvested annually from 2013 to 2015. The Basin cultivar consistently produced more seed than Timp (P < 0.05) , both on average and in each of the 3 years (Table 10; Peel et al. 2023).
Table 10. Seed production yields (lbs/ac) for Basin and Timp Utah sweetvetch cultivars grown for seed production in Millville, UT (Peel et al. 2023).
| Year | 2013 | 2014 | 2015 | Average |
| Basin (lbs/ac) | 633a¹ | 603a | 589a | 608a |
| Timp (lbs/ac) | 314b | 281b | 262b | 285b |
¹Values within a column with different letters are differ significantly (P < 0.05).
Nursery Practice
Nursery stock of Utah sweetvetch has been produced from seed (Johnson et al. 1989; Lapp et al. 2001). Plants can also be produced vegetatively from rhizomes of Utah sweetvetch accessions that produce rhizomes (Shaw 2004).
The following process was used to grow Utah sweetvetch from seed collected from 11 ecotypes growing in Montana, Idaho, Utah, and Colorado (Johnson et al. 1989). Seed was mechanically scarified (procedure not described), then inoculated with about 0.1 g Rhizobia/250 seeds (Hedysarum spec. 2 inoculum, Nitragin County, Milwaukee, WI). Inoculated seeds were placed in containers filled with equal parts sand, peat, and loam. Containers were placed in a greenhouse kept at 86 °F (30 °C) day and 59 °F (15 °C) night temperatures with no supplemental light (Johnson et al. 1989).
Nursery staff at Glacier National Park, Montana, used the following procedures to produce Utah sweetvetch plugs (Lapp et al. 2001). Seed was soaked in water for 24 hours, then sown in a 70% (6:1:1) peat, perlite, vermiculite and 30% sand mixture that filled 5.8 oz (172 ml) cone-tainers. Controlled release and micronutrient fertilizers were included with the growth media. Cone-tainers were put in the greenhouse at 70 to 77 °F (21-25 °C) day and 61 to 64 °F (16-18 °C) night temperatures. Media was kept moist during the germination period (about 3 weeks). Root development following germination was rapid, and shoots had four to five leaflets by week 10. Regular daily irrigation occurred until cone-tainers were thoroughly leached. Seedlings were fertilized (13-13-13 NPK at 100 ppm) until cone-tainers were root tight. Seedlings were moved to an outdoor nursery in mid-May and fertilized again (10-20-20 NPK at 200 ppm) in August and September. Irrigation was reduced in September and October, but seedlings were fully irrigated before winterizing. Plants not harvested in September were overwintered outdoors beneath insulating foam cover and snow for up to 5 months (Lapp et al. 2001).
Wildland Seeding And Planting
Utah sweetvetch has been used in many wildland restoration projects and in experimental wildland seeding and planting efforts (Plummer et al. 1968). Results were often mixed, but typically if Utah sweetvetch established successfully it persisted (Stevens and Monsen 2004), and several recommendations related to seeding and planting are available.
General Wildland Seeding Recommendations. Seeding in fall or early winter with seed that has been inoculated with an appropriate nitrogen-fixing Rhizobia species is recommended (Wasser 1982; Monsen and Stevens 2004; Stevens and Monsen 2004; Rosales 2014). Seed should be direct seeded at least 0.13 in (0.3 cm) but not more than 0.38 in (1 cm) deep in rows spaced 15 to 18 in (38-46 cm) (Stevens and Monsen 2004).
The pure stand seeding rate is 18 to 24 PLS lbs/ac (20-27 kg/ha) and should be adjusted according to the seeding mix (Ogle et al. 2003, 2011, 2019). For plans to deliver 1 seed/linear foot the rate is 1.3 lbs/ac (Monsen and Stevens 2004). In a seeding mix to revegetate sandy sites in mountain brushlands in Utah, Wasser (1982) suggests use of 0.5 lb/ac (0.6 kg/ha) of Utah sweetvetch if drill seeding and 1 lb/ac (1.1 kg/ha) if broadcast seeding. Because Utah sweetvetch is slow to establish, seeding in alternate rows from grasses and other rapidly growing species is recommended (Stevens 1987; Ogle et al. 2003; Stevens and Monsen 2004).
In the Intermountain West, Utah sweetvetch is used in revegetation efforts in Wyoming, basin, and mountain big sagebrush, pinyon-juniper, and ponderosa pine habitats where annual precipitation ranges from 10 to 36 in (254-914 mm) (Stevens and Monsen 2004). In Nevada, use of Utah sweetvetch in restoration is suitable in Major Land Resource Areas 24, 25, 28A, and 28B (Eldredge et al. 2013). In Glacier National Park, Montana, Utah sweetvetch was part of a seed mix used to restore fescue (Festuca spp.) grasslands (Majerus 1999). It is also recommended for pollinator habitat in the Black Hills coniferous forest ecoregion (SD and ne WY) and in the Plains, Palouse, and Dry Steppe province (MT, ND, SD, WY, CO, TX, ID, and WA) (Wojcik and Morandin 2008a, 2008b).
The species is persistent in rangeland communities with dry, often rocky, and well-drained sandy loam to silty clay loam soils where typical annual precipitation ranges from 12 to 18 in (305-457 mm) (Walker and Shaw 2005). In a study evaluating nodulation and acetylene reduction (nitrogen fixation) potential of 85 soil collections from 27 of the major soil associations in the northern Great Basin, nearly all supported indigenous Rhizobia spp. populations appropriate to nodulate Utah sweetvetch. The presence of juniper species at the collection site was associated with the highest acetylene reduction activity by Utah sweetvetch (Rumbaugh and Johnson 1984).
Utah sweetvetch is a long-lived and slow-growing species taking 3 to 4 years to establish, mature, and flower (Stevens 2004b). Based on research conducted on the restoration of big game habitat in Utah, Utah sweetvetch was rated good to very good in germination, initial establishment, growth rate, final establishment, and persistence (Plummer et al. 1968). It also rated good in ease of seeding, compatability with other species, seed production, forage yield, palatability, and tolerance to grazing (Stevens and Monsen 2004). Once established, Utah sweetvetch is persistent. It is drought tolerant, winter hardy, and a good choice for improving big game habitats, livestock range, and stabilizing disturbed or eroded sites (Stevens and Monsen 2004).
Grazing protection for at least 2 years is recommended at sites where Utah sweetvetch has been seeded. Because Utah sweetvetch is slow to establish but persistent and valuable once established, protection of newly seeded sites is important to protect the restoration investment (Jensen et al. 1999; Stevens 2004b; Stevens and Monsen 2004).
Wildland Seeding Study Results. In this section, studies that evaluated wildland seeding Utah sweetvetch are summarized. Because these studies vary widely by site condition, monitoring duration, and seeding method, they are best considered in context, which prevents synthesis of broad guidance or recommendations.
Utah sweetvetch established equally well on sites treated with two experimental surface disturbances in big sagebrush, pinyon-juniper, and mountain shrub communities in the Piceance Basin in Colorado (Sims and Redente 1974). Disturbance treatments were 1) vegetation removal by scraping and minimal topsoil loss or 2) vegetation removal followed by plowing to 8 to 12 in (20-30 cm) depths. Utah sweetvetch established on plowed and scraped sites. Emergence and survival were rated fair to good in year 1 and good in year 2. Utah sweetvetch performed best of all seeded forbs with germination as high as 87%. Seed was fall planted at 0.5 in (1.3 cm) deep at a rate of about 45 seeds/linear ft (150/linear m) (Sims and Redente 1974). In a follow-up study, Utah sweetvetch survival to year 6 or 7 was best at the highest elevation mountain brush site, but fair to poor at the sagebrush site. Utah sweetvetch did not survive to year 6 at the high-elevation pinyon-juniper site where precipitation was low, soils were shallow, and slopes were extreme (Redente 1980).
At the Curlew National Grassland near American Falls, Idaho, Timp Utah sweetvetch was still present 10 years after it was drill seeded on a site that burned in 2006 (Tilley et al. 2022). The site was plowed and packed in fall 2009, then herbicide-treated the following spring and summer. Drill seeding occurred on November 17, 2010, using a Tye grain drill and row spacings of 10 in (25 cm). Plots were mowed to a height of 10 in (25 cm) in late September 2011 to prevent seed set of weed species. Plots received no other treatments and no supplemental irrigation. Initial establishment (2011) of Utah sweetvetch was high (0.7 plants/ft2 [7.6 /m2]), but density declined from 2011 to 2012 (~ 0.1/ft2 [1/m2] in 2013). In 2020, Timp Utah sweetvetch was still present on the site (0.13 plants/ft2 [1.4/m2]). Drought conditions in 2012 and reestablishment of crested wheatgrass (Agropyron cristatum) and bulbous bluegrass (Poa bulbosa) were likely important in the decreased abundance of seeded species (Tilley et al. 2022).
A seeding study in Canada evaluated establishment and short-term survival of Utah sweetvetch at two prairie sites (Quilichini et al. 2021). Utah sweetvetch had better establishment and survival at the moist, nutrient-rich site (Saskatoon, SK), but establishment was improved with higher seeding rates at the dry, nutrient-poor site (Swift Current, SK). The study also evaluated seeding of legume mixes that included Utah sweetvetch and other legume species with meadow brome (Bromus riparius). Plots were treated with glyphosate, harrowed, and packed prior to seeding in mid-May 2016 using a 6-row disc seeder. Seed was planted 0.5 in (1.3 cm) deep in rows spaced 1 ft (0.3 m) apart. Plots were hand weeded in the establishment year and mowed multiple times in the first growing season to control weeds. Cover of Utah sweetvetch at the moist, more nutrient rich site was 26% in the establishment year (2016) and 42% in 2018. Cover was 4% in 2016 and 0% in 2018 at the drier, nutrient-poor site. No data was collected for individual legume species in mixture seedings, but as a group native legumes contributed little (<10%) cover and forage at either site (Quilichini et al. 2021). In a companion study, seedling establishment by seeding rate was evaluated for monoculture stands at the dry, low-nutrient site (Table 11). Seed was scarified and seeded on May 18, 2018, using the same method describe above at seeding rates of 30, 61, and 91 PLS/ft (100, 200, 300 PLS/m) of row. Increased seeding rates increased establishment (Quilichini et al. 2021).
Table 11. Count of Utah sweetvetch seedlings (count/ft2) of Utah sweetvetch at Swift Current, SK with increased seeding rates at a dry, low-nutrient site seeded on May 18, 2018 (Quilichini et al. 2021).
| Seeding rate (PLS/linear ft) | 30 | 61 | 91 |
| Seedling density July 2018 |
0.7a¹ | 0.7a | 0.5a |
| Seedling density September 2018 |
0.5a | 1.1b | 1.1b |
¹Values in a row with different letters are significantly different (P < 0.05).
Utah sweetvetch establishment was considered successful in the revegetation of gravel-dominated pits and pads in Churchill, northern Manitoba (Rausch and Kershaw 2007). Revegetation treatments were implemented on sites abandoned 5 to 42 years earlier. Treatments included snow fencing, microrelief alteration, and seeding. Seed was mixed with the top 1 in (2.5 cm) of gravel, peat, and fertilizer. Although density and cover of Utah sweetvetch were reduced by almost 50% between year 1 and 2, researchers anticipated that seeding earlier (seeding done in early July in this study), using a better adapted seed source (seed used was from northern Alberta in this study), and increasing the seeding rate would improve future success with Utah sweetvetch (Rausch and Kershaw 2007).
Utah sweetvetch was one of 14 legume species collected and evaluated for use in reclamation in Alberta (Smreciu 1995). Seed was collected from 41 foothills and mountain sites in Alberta. The seed was grown in a greenhouse then outplanted in a field nursery (Vegreville) in late spring and observed for 3 seasons. Although mortality was high over the first winter, it was low in following years as both vigor and cover of Utah sweetvetch increased. Utah sweetvetch produced some seed in the first year, but seed production was low likely because of poor pollination (Smreciu 1995).
Native forb establishment was poor in Wyoming big sagebrush vegetation where four seeding techniques were compared in Toole County, Utah (mean annual ppt: 13 in [330 mm]) (Gunnell and Summers 2016). Plots were either drill seeded using the Truax Rough Rider or broadcast seeded, followed by Lawson aerator, Dixie harrow, or Ely Chain treatments. There were nine forb species in the seeded mix (seeding rate for Utah sweetvetch: 0.6 PLS/ft2 [6.5 PLS/m2]). For any technique, total survival of all forbs was less than 6%. Establishment of Utah sweetvetch was greatest on chain treatments in year 1, but survival declined in subsequent years. Drill seeded plots had the lowest establishment and survival in all years (Gunnell and Summers 2016).
Use of row cover to establish islands of native forbs in big sagebrush communities was field tested at four sites in Utah (three in Wyoming big sagebrush [Ecoregions 13 and 19] and one in mountain big sagebrush [Ecoregion 20]) (Landeen et al. 2021). Seedling emergence was often greater for Timp with row cover, but differences did not persist beyond the first year (Table 12). Timp Utah sweetvetch emerged at all sites and persisted in small quantities (<0.05 plants/ft2 [0.5 plants/m2]) in covered and noncovered plots at all sites. The seeding rate for Timp was 2.9 PLS/ft2 (31.7 PLS/m2), which is higher than typical. Sites were cleared of vegetation using a Dixie Harrow, the seed was broadcast using a hand-crank seeder followed by a Brillion imprinter and lightweight chain to incorporate the seed into the soil. In control plots (unseeded with and without row cover), undesirable species increased with row cover treatments suggesting that increases in competition could accompany row cover treatments. Seed source may have also been a reason for limited establishment and persistence in this study, however this does not account for the overall failure of seed island establishment (Landeen et al. 2021).
Table 12. Mean density of Utah sweet vetch with and without row cover (N-sulate) (DeWitt Co, Sikeston, MO) following seeding at four sites in Utah (Landeen et al. 2021).
| Year | ARTRW¹ (Ecoregion 19) Fountain Green |
ARTRV (Ecoregion 20) Gordon Creek |
ARTRW (Ecoregion 13) Hatch Ranch |
ARTRW (Ecoregion 13) Lookout Pass |
All Sites | |||||
| C² | NC | C | NC | C | NC | C | NC | C | NC | |
| 1 | 1.46 | 1.07 | 2.91³ | 1.27 | 4.05 | 1.12 | 1.98 | 0.60 | 2.60 | 1.01 |
| 2 | 0 | 0.02 | 0.51 | 0.43 | 0.14 | 0.16 | 0.12 | 0.06 | 0.19 | 0.17 |
| 5 | 0 | 0 | 0.03 | 0.07 | 0.33 | 0.07 | 0 | 0 | 0.09 | 0.03 |
¹ARTRW = Artemisia tridentata subsp. wyomingensis, ARTRV = A. t. subsp. vaseyana.
²C = covered, NC = not covered.
³Bold values within a site show significantly different plant densities between covered and noncovered plots (P < 0.001).
Mine Remediation Seeding and Planting Study Results. Studies of seeding Utah sweetvetch on abandoned mine sites showed mixed results but establishment was improved when seedlings were used.
Utah sweetvetch established on amended and unamended mine spoils where it was seeded and where it was not purposely seeded in southeastern British Columbia (Smyth 1997a). The site was a 3-year-old coal mine spoil at an alpine tundra-spruce fir transition zone. Vegetation establishment was evaluated on seeded and unseeded plots with unamended spoils or spoils amended with native soil. Unamended spoils were primarily siltstone and carbonaceous shale. Amended spoils were covered with 2 in (5 cm) of native soil with vegetation and organic material removed and fertilizer added (171 lbs/ac [192 kg/ha] N, 17 lbs/ac [19 kg/ha] P and K). Seeds were collected locally and stored at 36 to 41 °F (2-5 °C) until used. In germination tests for seeded species (68 °F [20 °C], 12-hr/12-hr light-dark cycle), Utah sweetvetch germination was 84%. Utah sweetvetch seed was scarified before germination testing and prior to seeding at the reclamation site. Scarification was accomplished with a Hoffman Model SC-50 Scarifier lined with 80 grit garnet paper and operated at 60 to 80 PSI for 1 to 5 minutes. Seed was also inoculated with species-specific peat-based Rhizobium inoculum from nodules excised the previous summer. Spoil plots were seeded in May 1985 and establishment evaluated from 1986 to 1990. Utah sweetvetch established with low cover (<1%) on all but the unseeded unamended spoil plots. It was present in all post-seeding years and increased in abundance in years 4 and 5 on seeded amended soil plots. On seeded unamended spoil plots it was present for 3 years. It was present in post-seeding years 2 to 5 on unseeded amended soil plots where establishment was attributed to seed dispersal of 3 to 7 ft (1-2 m) from adjacent treatments (Smyth 1997a).
Utah sweetvetch establishment was poor on a recontoured surface mine site near Kemmerer, Wyoming (Hatton and West 1987). The site was recontoured with the top 6 in (15 cm) of soil replaced in fall 1981. Seeding of Utah sweetvetch (10 seeds/plot) and western wheatgrass (90 seeds/plot) occurred in spring 1982 in 0.25 m2 circular plots. Vegetation establishment was monitored until 1985. The study provided no other details about the seeding procedure or vegetation persistence (Hatton and West 1987). Utah sweetvetch failed to establish on retorted shale waste in northwest Colorado. The sites had 6 in (15 cm) of soil layered on top of the retorted shale prior to being seeded (Gerschefske-Kitt and Sabey 1987).
On unamended coal mine soils in British Columbia’s subalpine regions, Utah sweetvetch established but persistence was limited (Table 13, Smyth 1997b). Transplants were grown from a single accession collected adjacent to the reclamation site. Seedlings were 4 months old at the time of outplanting. Damage and mortality from bighorn sheep use were extensive (Smyth 1997b).
Table 13. Survival and growth of Utah sweetvetch outplants on an unamended coal mine spoil in British Columbia. Seedlings were spaced 10 in (25 cm) apart (Smyth 1997b).
| Site | Post-treatment year | Number surviving | % flowering | % seedheads | % eaten | Mean height (in) | Mean width (in) |
| Line Creek | 1 | 100 | 0 | 0 | 100 | 2.7 | 2.6 |
| 5 | 6 | 0 | 0 | 100 | 2.9 | 3.5 | |
| Quinette | 1 | 100 | 0 | 0 | 10 | 2.7 | 3.3 |
| 5 | 52 | 75 | 60 | 25 | 3.0 | 5.5 |
The effects of adding mycorrhizal fungi and Rhizobia (pure inoculum) on the growth of seedlings that were planted on a surface coal mine were evaluated near Kemmerer, Wyoming (Carpenter and Allen 1988). The site was mined in the late 1970s then backfilled and regraded in 1979 with no topsoil applied. Utah sweetvetch seed was collected in July 1982 from Rich County, Utah, about 37 mi (60 km) from the mine sites. Seed was grown in plastic tube-paks (Native Plants Inc., SLC, UT) filled with soils collected from the mine spoil with the following treatments: Rhizobia (seeds coated with Rhizobia), vesicular arbuscular (VA) mycorrhizal (seeds coated with topsoil), Rhizobia + VA mycorrhiza (coated with rhizobia and topsoil). Each tube had 0.8 in (2 cm) of topsoil added. Seed was grown in a heated glasshouse over the winter and transplanted in mid-June 1983. Plants receiving both inoculations had greatest survival to August 1985 (Table 14, Carpenter and Allen 1988).
Table 14. Biomass production and early reproductive output of Utah sweetvetch transplants receiving different seed treatments (vesicular arbuscular [VA] mycorrhizal fungi, Rhizobia, fungi + Rhizobia, or none) (Carpenter and Allen 1988).
| Treatment | Control | +VA mycorrhiza | +Rhizobia | + VA and Rhizobia |
| Above ground biomass (g) in 1984 | 0.77a¹ (n=4) | 0.77a (n=5) | 1.13b (n=3) | 1.79c (n=12) |
| Above ground biomass (g) in 1985 | 2.24a (n=34) | 1.92a (n=31) | 2.16a (n=17) | 5.09b (n=23) |
¹Values within a row followed by different letters are significantly different (P < 0.05).
Other Restoration Management. Some studies suggest Utah sweetvetch is competitive with spotted knapweed (Centaurea maculosa) and thus potentially useful in restoration efforts where spotted knapweed is a problem (Perry et al. 2005). A study that evaluated Utah sweetvetch’s resistance to catechin, a potent phytotoxin exuded from roots of spotted knapweed, rated Utah sweetvetch as moderately resistant. Germination, root and shoot growth, and seedling mortality of Utah sweetvetch were not severely impacted at catechin concentrations of 0 to 4.0 mg/mL (Perry et al. 2005).
In greenhouse competition experiments, Utah sweetvetch was grown with bluebunch wheatgrass (Pseudoroegneria spicata) and spotted knapweed (Jacobs and Sheley 1999). Based on these experiments, researchers found that growth characteristics of Utah sweetvetch were more similar to those of spotted knapweed than to bluebunch wheatgrass, and that Utah sweetvetch competed more directly with spotted knapweed. These findings support the hypothesis that tap-rooted forbs growing with bunch grasses increases niche occupation and may be effective in minimizing invasion by tap-rooted weeds. Greenhouse conditions and seedling growth of Utah sweetvetch are described in the earlier Seedling Ecology section (Jacobs and Sheley 1999).
Although the above studies suggest Utah sweetvetch is competitive with spotted knapweed, a study of the Going-to-the-Sun highway in Glacier National Park, Montana, suggested otherwise. Tyser (1992) found that Utah sweetvetch was absent from spotted knapweed- and timothy (Phleum pratense)-invaded vegetation but its cover was 0.5% in native fescue-dominated vegetation (Tyser 1992).
Acknowledgements
Funding for Western Forbs: Biology, Ecology, and Use in Restoration was provided by the USDI BLM Great Basin Native Plant Materials Ecoregional Program through the Great Basin Fire Science Exchange. Great thanks to the chapter reviewers: Noelle Orloff, Montana State University Extension and Byron Love, Agricultural Research Service.
This research was supported in part by the USDA Forest Service, Rocky Mountain Research Station. The findings and conclusions in this publication are those of the authors and should not be construed to represent any official USDA or U.S. Government determination or policy.
Literature Cited
Arno, S.F.; Simmerman, D.G.; Keane, R.E. 1985. Forest succession on four habitat types in western Montana. Gen. Tech. Report INT-177. Ogden, UT: U.S. Department of Agriculture, Forest Service, Intermountain Forest and Range Experiment Station. 74 p.
Association of Official Seed Analysts [AOSA]. 2010. AOSA/SCST Tetrazolium testing handbook. Contribution No. 29. Lincoln, NE: Association of Official Seed Analysts.
Association of Official Seed Analysts [AOSA]. 2023. AOSA rules for testing seeds. Vol. 1. Principles and procedures. Washington, DC: Association of Official Seed Analysts.
Bamberg, S.A.; Major, J. 1968. Ecology of the vegetation and soils associated with calcareous parent materials in three alpine regions of Montana. Ecological Monographs. 38(2): 12-167.
Barneby, R.C. 1989. Intermountain Flora Volume 3, Part B: Fabales. In: Cronquist, A.; Holmgren, A.H.; Holmgren, N.H.; Reveal, J.L.; Holmgren, P.K., eds. Intermountain flora: Vascular plants of the Intermountain West, U.S.A. Bronx, NY: The New York Botanical Garden. 279 p.
Basey, A.C.; Fant, J.B.; Kramer, A.T. 2015. Producing native plant materials for restoration: 10 rules to collect and maintain genetic diversity. Native Plants Journal. 16(1): 37-53.
Belcher, E. 1985. Handbook on seeds of browse-shrubs and forbs. Tech. Pub. No. R8-TP8. Atlanta, GA: U.S. Department of Agriculture, Forest Service, Southern Region. 246 p.
Birks, H.J.B. 1980. The present flora and vegetation of the moraines of the Klutlan Glacier, Yukon Territory, Canada: A study in plant succession. Quaternary Research. 14: 60-86.
Bishop, S.C.; Chapin, F.S. 1989. Patterns of natural revegetation on abandoned gravel pads in arctic Alaska. Journal of Applied Ecology. 26(3): 1073-1081.
Blundon, D.J.; Dale, M.R.T. 1990. Dinitrogen fixation (Acetylene reduction) in primary succession near Mount Robson, British Columbia, Canada. Arctic and Alpine Research. 22(3): 255-263.
Blundon, D.J.; Macisaac, D.A.; Dale, M.R.T. 1993. Nucleation during primary succession in the Canadian Rockies. Canadian Journal of Botany. 71(8): 1093-1096.
Boukheloua, J. 1990. Germination, growth, nitrogen status, and salt tolerance of Hedysarum boreale. Fort Collins, CO: Colorado State University. Thesis. 60 p.
Bower, A.D.; St. Clair, J.B.; Erickson, V. 2014. Generalized provisional seed zones for native plants. Ecological Applications. 24(5): 913-919.
Bushman, B.S.; Larson, S.R.; Peel, M.D.; Pfrender, M.E. 2007. Population structure and genetic diversity in north American Hedysarum boreale Nutt. Crop Science. 47(3): 1281-1288.
Cane, J.H. 2005. Pollinator and seed predator studies. In: Shaw, N.L.; Pellant, M., eds. Great Basin Native Plant Project: 2006 Progress Report. Boise, ID: U.S. Department of Agriculture, Rocky Mountain Research Station: 23-26.
Carpenter, A.T.; Allen, M.F. 1988. Responses of Hedysarum boreale Nutt. to mycorrhizas and rhizobium – Plant and soil nutrient changes in a disturbed shrub-steppe. New Phytologist. 109(1): 125-132.
Catling, P.M.; Kostiuk, B.; Thompson, D. 2015. Horse ranching increases biodiversity in a foothills parkland prairie in Northern Kananaskis Country, western Alberta. The Canadian Field-Naturalist. 129(1): 15-23.
Consortium of Pacific Northwest Herbaria [CPNWH]. 2020. Seattle, WA: University of Washington Herbarium, Burke Museum of Natural History and Culture. http://www.pnwherbaria.org/index.php
Craighead, J.J.; Craighead, F.C. Jr.; Davis, R.J. 1963. A field guide to Rocky Mountain wildflowers from northern Arizona and New Mexico to British Columbia. Boston, MA: Houghton Mifflin Company. 277 p.
Discover Life. 2024. Sam Houston State University. Available: https://www.discoverlife.org/
Doak, D.F.; Loso, M.G. 2003. Effects of grizzly bear digging on alpine plant community structure. Arctic, Antarctic, and Alpine Research. 35(4): 421-428.
Dorn, R.D.; Dorn, J.L. 2007. Growing native plants of the Rocky Mountain area. Morrisville, NC: Lulu.com. 260 p.
Dreesen, D.R.; Harrington, J.T. 1997. Propagation of native plants for restoration projects in the southwestern U.S.–preliminary investigations. In: Landis, T.D.; Thompson, J.R., tech. coords. National proceedings: Forest and Conservation Nursery Associations–1997; Regeneration, reforestation, restoration: The seedling is the key. 1997 August 19-21; Boise, ID. Portland, OR: U.S. Department of Agriculture, Forest Service, Pacific Northwest Research Station: 77-88.
Duan, L.; Wen, J.; Yang, X.; Liu, P.L.; Arslan, E.; Ertugrul, K.; Chang, Z.Y. 2015. Phylogeny of Hedysarum and tribe Hedysareae (Leguminosae: Papilionoideae) inferred from sequence data of ITS, matK, trnL-F and psbA-trnH. Taxon. 64(1): 49-64.
EcoRestore Portal: Restore Arizona’s Native Plants [EcoRestore AZ]. 2024. University of Arizona. Available: https://ecorestore.arizona.edu/
Eddleman, L.E. 1977. Indigenous plants of southeastern Montana. I. Viability and suitability for reclamation in the Fort Union Basin. Spec. Pub. 4. Missoula, MT: University of Montana, Montana Forest and Conservation Experiment Station. 122 p.
Eldredge, E.; Novak-Echenique, P.; Heater, T.; Mulder, A.; Jasmine, J. 2013. Plants for pollinator habitat in Nevada. Tech. Note NV 57. Reno, NV: U.S. Department of Agriculture, Natural Resources Conservation Service. 65 p.
European Native Seed Conservation Network [ENSCONET]. 2009. ENSCONET seed collecting manual for wild species. Edition 1: 32 p.
Ford, T.M.J. 1988. Establishment, nitrogen fixation, and nutritive value of Utah sweetvetch (Hedysarum boreale Nutt.). Logan, UT: Utah State University. Thesis. 72 p.
Ford, T.M.J.; Johnson, D.A.; Rumbaugh, M.D.; Richardson, B.Z. 1989. Sweetvetch: A native legume for rangeland revegetation. Utah Science. 50(2): 73-77.
Gerschefske-Kitt, D.G.; Sabey, B.R. 1987. Vegetative stabilization of retorted oil shale on a steep, south-facing slope in Northwestern Colorado. Landscape and Urban Planning. 14: 497-506.
Glenny, W.; Runyon, J.; Burkle, L. 2022. Assessing pollinator friendliness of plants and designing mixes to restore habitat for bees. Gen Tech. Rep. RMRS-GTR-429. Fort Collins, CO: USDA Forest Service, Rocky Mountain Research Station. 52 p.
Goodrich, S.; Neese, E. 1986. Uinta Basin flora. Ogden, UT: U.S. Department of Agriculture, Forest Service, Intermountain Region. 320 p.
Gunnell, K.; Summers, D. 2016. Native forb increase and research at the Great Basin Research Center. In: Kilkenny, F.; Edwards, F.; Malcomb, A., eds. Great Basin Native Plant Project: 2015 Progress Report. Boise, ID: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station: 96-108.
Hammon, R. 2007. Insect pests of selected grass and forb species in the Great Basin. In: Shaw, N.; Pellant, M., eds. Great Basin Native Plant Selection and Increase Project: 2006 Progress Report. Boise, ID: U.S. Department of Agriculture, Rocky Mountain Research Station: 93-94.
Hatton, T.; West, N. 1987. Early seral trends in plant community diversity on a recontoured surface mine. Vegetatio. 73(1): 21-29.
Hermann, F. 1966. Notes on western range forbs: Cruciferae through Compositae. Agric. Handb. 293. Washington, DC: U.S. Department of Agriculture, Forest Service. 365 p.
Hess, J. 2011. Greater sage-grouse (Centrocercus urophasianus) habitat response to mowing and prescribed burning Wyoming big sagebrush and the influence of disturbance factors on lek persistence in the Bighorn Basin, Wyoming. Laramie, WY: University of Wyoming. Thesis. 152 p.
Hitchcock, C.L.; Cronquist, A. 2018. Flora of the Pacific Northwest: An illustrated manual. Second Ed. Giblin, D.E.; Legler, B.S.; Zika, P.F.; Olmstead, R.G., eds. Seattle, WA: University of Washington Press. 882 p.
Hollingsworth, T.N.; Lloyd, A.H.; Nossov, D.R.; Ruess, R.W.; Charlton, B.A.; Kielland, K. 2010. Twenty-five years of vegetation change along a putative successional chronosequence on the Tanana River, Alaska. Canadian Journal of Forest Research. 40(7): 1273-1287.
ITIS Database. 2024. Integrated Taxonomic Information System. Available: http://www.itis.gov/index.html
Jacobs, J.S.; Sheley, R.L. 1999. Competition and niche partitioning among Pseudoroegneria spicata, Hedysarum boreale, and Centaurea maculosa. The Great Basin Naturalist. 59(2): 175-181.
Jensen, K.; Horton, H.; Reed, R.; Whitesides, R.; USDA-ARS-FRRL. 1999. Intermountain planting guide. Logan, UT: U.S. Department of Agriculture, Agricultural Research Service, Range Research Lab and Utah State University Extension. 104 p.
Johnson, C.D. 1990. Confirmation of Hedysarum boreale Nuttall (Leguminosae) as a host plant for Acanthoscelides fraterculus (Horn) (Coleoptera, Bruchidae). Pan-Pacific Entomologist. 66(2): 175-176.
Johnson, D.A.; Ford, T.M.J.; Rumbaugh, M.D.; Richardson, B.Z. 1989. Morphological and physiological variation among ecotypes of sweetvetch (Hedysarum boreale Nutt.). Journal of Range Management. 42(6): 496-501.
Jorgensen, K.R.; Stevens, R. 2004. Seed collection, cleaning, and storage. In: Monsen, S.B.; Stevens, R.; Shaw, N.L., comps. Restoring western ranges and wildlands, vol. 3. Gen. Tech. Rep. RMRS-GTR-136-vol-3. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station: 699-716.
Jorgensen, K.R.; Wilson, G.R. 2004. Seed germination. In: Monsen, S.B.; Stevens, R.; Shaw, N.L., comps. Restoring western ranges and wildlands, vol. 3. Gen. Tech. Rep. RMRS-GTR-136-vol-3. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station: 723-732.
Kitchen, S.G. 1994. Perennial forb life-history strategies on semiarid rangelands: Implications for revegetation. In: Monsen, S.B.; Kitchen, S.G., comps. Proceedings – Ecology and management of annual rangelands; 1992 May 18-22; Boise, ID. Gen. Tech. Rep INT-GTR-313. Ogden, UT: U.S. Department of Agriculture, Forest Service, Intermountain Research Station: 342-346.
Kudo, G.; Harder, L.D. 2005. Floral and inflorescence effects on variation in pollen removal and seed production among six legume species. Functional Ecology. 19(2): 245-254.
Lady Bird Johnson Wildflower Center [LBJWC]. 2013. Hedysarum boreale Nutt. Native Plant Database. Austin, TX: Lady Bird Johnson Wildflower Center. https://www.wildflower.org/plants/result.php?id_plant=HEBO [Accessed 2024 December 17].
Landeen, M.; Jones, C.; Jensen, S.; Whittaker, A.; Summers, D.D.; Eggett, D.; Petersen, S.L. 2021. Establishing seed islands for native forb species on rangelands using N-Sulate ground cover fabric. Native Plants Journal. 22(1): 51-63.
Lapp, J.; Evans, J.; Wick, D. 2001. Propagation protocol for production of container (plug) Hedysarum boreale Nutt. Plants 172 ml conetainers. Native Plant Network. U.S. Department of Agriculture, Forest Service, National Center for Reforestation, Nurseries, and Genetic Resources. http://npn.rngr.net/propagation/protocols [Accessed 2024 December 17].
Lesica, P. 2012. Manual of Montana vascular plants. Fort Worth, TX: Botanical Research Institute of Texas (BRIT) Press. 779 p.
Löve, A. 1982. IOPB chromosome number reports LXXV. Taxon. 31(2): 342-368.
Luna, T.; Mousseaux, M.R.; Dumroese, R.K. 2018. Common native forbs of the northern Great Basin important for greater sage-grouse. Gen. Tech. Rep. RMRS-GTR-387. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station; Portland, OR: U.S. Department of the Interior, Bureau of Land Management, Oregon-Washington Region. 76 p.
MacKinnon, A.; Kershaw, L.; Arnason, J.; Owen, P.; Karst, A.; Hamersley Chambers, F. 2014. Edible and medicinal plants of Canada. Edmonton, Alberta: Partners Publishing and Lone Pine Publishing. 448 p.
Majerus, M. 1999. Collection and production of indigenous plant material for national park restoration. In: Holzworth, L.K.; Brown, R.W., comps. Revegetation with native species: Proceedings, 1997 Society for Ecological Restoration annual meeting; 1997 November 12-15; Fort Lauderdale, FL. Proc. RMRS-P-8. Ogden, UT: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station: 17-21.
McGuire, A.D. 1993. Interactions for pollination between two synchronously blooming Hedysarum species (Fabaceae) in Alaska. American Journal of Botany. 80(2): 147-152.
Mee, W.; Barnes, J.; Kjelgren, R.; Sutton, R.; Cerny, T.; Johnson, C. 2003. Water wise: Native plants for Intermountain landscapes. Logan, UT: Utah State University Press. 220 p.
Meyer, S.E. 2005. Intermountain Native Plant Growers Association: A nonprofit trade organization promoting landscape use of native plants. Native Plants Journal. 6(2): 104-107.
Meyer, S.E.; Kjelgren, R.K.; Morrison, D.G.; Varga, W.A. 2009. Chapter 7: The plant palette. In: Landscaping on the new frontier: Waterwise design for the Intermountain West. Logan, UT: Utah State University Press: 159-226.
Moerman, D. 2003. Native American ethnobotany: A database of foods, drugs, dyes, and fibers of Native American peoples, derived from plants. Dearborn, MI: University of Michigan. http://naeb.brit.org
Monsen, S.B.; Stevens, R. 2004. Seedbed preparation and seeding practices. In: Monsen, S.B.; Stevens, R.; Shaw, N.L., comps. Restoring western ranges and wildlands, vol. 1. Gen. Tech. Rep. RMRS-GTR-136-vol-1. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station: 121-154.
Northstrom, T.E. 1969. A revision of the Hedysarum boreale complex. Provo, UT: Brigham Young University. Dissertation. 187 p.
Ogle, D.; Pavek, P.; Fleenor, R.; Stannard, M.; Dring, T.; Cane, J.; Fullen, K.; St. John, L.; Tilley, D. 2011. Plants for pollinators in the Inland Northwest. Tech. Note 2B. Boise, ID: U.S. Department of Agriculture, Natural Resources Conservation Service. 65 p.
Ogle, D.; St. John, L.; Stannard, M.; Holzworth, L. 2012. Conservation plant species for the Intermountain West. Plant Material Technical Note 24. Boise, ID: U.S. Department of Agriculture, Natural Resources Conservation Service. 57 p.
Ogle, D.; Tilley, D.; Cane, J.; St. John, L.; Fullen, K.; Stannard, M.; Pavek, P. 2019. Plants for pollinators in the Intermountain West. Plant Materials Tech. Note 2A. Boise, ID: U.S. Department of Agriculture, Natural Resources Conservation Service. 40 p.
Omernik, J.M. 1987. Ecoregions of the conterminous United States. Map (scale 1:7,500,000). Annals of the Association of American Geographers. 77(1): 118-125.
Parker, K.G. 1980. Some important Utah range plants. EC 383. Logan, UT: Utah State University Cooperative Extension Service. 174 p.
Pavek, P.; Erhardt, B.; Heekin, T.; Old, R. 2012. Forb seedling identification guide for the Inland Northwest: Native, introduced, invasive, and noxious species. Pullman, WA: U.S. Department of Agriculture, Natural Resources Conservation Service, Pullman Plant Materials Center. 144 p.
Peck, C.H. 1881. New species of fungi. Botanical Gazette. 6(10): 274-279.
Peel, M.D.; Ransom, C.V.; Mott, I.W.; Waldron, B.L. 2015. Natural glyphosate tolerance in sweetvetch Hedysarum boreale. Crop Science. 55(5): 2368-2376.
Peel, M.D.; Waldron, B.L.; Walker, S. 2011. A growth stage and heat-unit model for Hedysarum boreale. Native Plants Journal. 12(1): 4-12.
Peel, M.D.; Walker, S.; Waldron, B.L. 2023. Notice of release of ‘Basin’ Utah sweetvetch. Native Plants Journal. 24(3): 273-280.
Perry, L.G.; Johnson, C.; Alford, E.R.; Vivanco, J.M.; Paschke, M.W. 2005. Screening of grassland plants for restoration after spotted knapweed invasion. Restoration Ecology. 13(4): 725-735.
Pigeon, K.E.; Cote, S.D.; Stenhouse, G.B. 2016. Assessing den selection and den characteristics of grizzly bears. The Journal of Wildlife Management. 80(5): 884-893.
Plant Conservation Alliance [PCA]. 2015. National seed strategy for rehabilitation and restoration 2015-2020. Washington, DC: U.S. Department of the Interior, Bureau of Land Management. 52 p.
Plants for a Future [PFAF]. 2024. Available: https://pfaf.org/user/Plant.aspx
Plummer, A.P.; Christensen, D.R.; Monsen, S.B. 1968. Restoring big-game range in Utah. Pub. 68-3. Salt Lake City, UT: Utah Division of Fish and Game. 183 p.
Quilichini, S.M.; Schellenberg, M.P.; Biligetu, B. 2021. Introduction of four native legumes of the Canadian Prairies in monoculture and in mixtures with meadow bromegrass (Bromus riparius). Canadian Journal of Plant Science. 101(5): 714-729.
Rausch, J.; Kershaw, G.P. 2007. Short-term revegetation performance on gravel-dominated, human-induced disturbances, Churchill, Manitoba, Canada. Arctic, Antarctic, and Alpine Research. 39(1): 16-24.
Redente, E.F. 1980. Autecology of Hedysarum boreale. Fort Collins, CO: Colorado State University. Dissertation. 73 p.
Redente, E.F. 1982. Sweetvetch seed germination. Journal of Range Management. 35(4): 469-472.
Redente, E.F.; Reeves, F.B. 1981. Interactions between vesicular-arbuscular mycorrhiza and Rhizobium and their effect on sweetvetch growth. Soil Science. 132(6): 410-415.
Richardson, B.; Kilkenny, F.; St. Clair, B.; Stevenson-Molnar, N. 2020. Climate Smart Restoration Tool. https://climaterestorationtool.org/csrt/
Robinson, G.S.; Ackery, P.R.; Kitching, I.; Beccaloni, G.W.; Hernández, L.M. 2025. HOSTS: A Database of the World’s Lepidopteran Hostplants [Data set]. Natural History Museum. https://doi.org/10.5519/havt50xw
Robus, M.A. 1981. Muskox habitat and use patterns in northeastern Alaska. Fairbanks, AK: University of Alaska. Thesis. 116 p.
Rosales, M. 2006. ‘Timp’ Utah sweetvetch release brochure. Meeker, CO: U.S. Department of Agriculture, Natural Resources Conservation Service, Upper Colorado Environmental Plant Center. 2 p.
Rosales, M. 2014. Plant guide: Utah sweetvetch (Hedysarum boreale Nutt.). Meeker, CO: U.S. Department of Agriculture, Natural Resources Conservation Service, Upper Colorado Environmental Plant Center. 4 p.
Ross, C.J. 2020. Hedysarum (Sweetvetch). In: Meyers, S.C.; Jaster, T.; Mitchell, K.E.; Harvey, T.; Hardison, L.K., eds. Flora of Oregon, Vol. 2: Dicots A-F. Fort Worth, TX: Botanical Research Institute of Texas (BRIT) Press: 682.
Rouse, W.R.; Kershaw, K.A. 1973. Studies on lichen-dominated systems. VI. Interrelations of vegetation and soil moisture in the Hudson Bay Lowlands. Canadian Journal of Botany. 51(7): 1309-1317.
Rumbaugh, M.D. 1983. Legumes–their use in wildland plantings. In: Monsen, S.B.; Shaw, N., comps. Managing Intermountain rangelands–Improvement of range and wildlife habitats: Proceedings of symposia. Gen. Tech. Rep. INT-157. 1981 September 15-17, Twin Falls, ID; 1982 June 22-24, Elko, NV. Ogden, UT: U.S. Department of Agriculture, Forest Service, Intermountain Forest and Range Experiment Station: 115-122.
Rumbaugh, M.D.; Johnson, D.A. 1984. Nodulation and acetylene-reduction by two legumes with rhizobia indigenous to northern Great Basin soils. Great Basin Naturalist. 44(1): 151-158.
SEINet – Regional Networks of North American Herbaria Steering Committee [SEINet]. 2020. SEINet Regional Networks of North American Herbaria. https://Symbiota.org/docs/seinet
Shaw, N.L. 2004. Production and use of planting stock. In: Monsen, S.B.; Stevens, R.; Shaw, N.L., comps. Restoring western ranges and wildlands, vol. 3. Gen. Tech. Rep. RMRS-GTR-136-vol-3. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station. p. 745-768.
Shaw, R.J. 1995. Utah wildflowers: A field guide to northern and central mountains and valleys. Logan, UT: Utah State University Press. 218 p.
Silva, S.; Ayers, T. 2016. Plant endemism on Mancos Shale barrens. Natural Areas Journal 36(2): 166-176.
Sims, P.L.; Redente, E.F. 1974. Revegetation of disturbed surface soils in various vegetation ecosystems of the Piceance Basin. Tech. Rep. 5. Fort Collins, CO: Colorado State University, Environmental Resources Center. 157 p.
Smreciu, A. 1995. Selection of native legume species for reclamation in the Rocky Mountains. In: Proceedings of the 19th Annual British Columbia Mine Reclamation Symposium. 1995 June 19-23; Dawson Creek, BC: 14-23.
Smyth, C.R. 1997a. Early succession patterns with a native species seed mix on amended and unamended coal mine spoil in the Rocky Mountains of southeastern British Columbia, Canada. Arctic and Alpine Research. 29(2): 184-195.
Smyth, C.R. 1997b. Native legume transplant survivorship and subsequent seedling recruitment on unamended coal mine soils in the Canadian Rocky Mountains. Mountain Research and Development. 17(2): 145-157.
Society for Ecological Restoration; International Network for Seed Based Restoration; Royal Botanic Gardens [SER, INSR, RBGK]. 2024. Seed Information Database (SID). Available from: https://ser-sid.org/
Spellman, B.T.; Wurtz, T.L. 2011. Invasive sweetclover (Melilotus alba) impacts recruitment along floodplains of interior Alaska. Biological Invasions. 13(8): 1779-1790.
Stevens, O.A. 1957. Weights of seeds and numbers per plant. Weeds. 5(1): 46-55.
Stevens, R. 1987. Thirty years of pinyon-juniper big game habitat improvement projects: What have we learned? In: Everett, R.L., comp. Proceedings–pinyon-juniper conference. 1986 January 13-16; Reno, NV. Gen. Tech. Rep. INT-GTR-215. Ogden, UT: U.S. Department of Agriculture, Forest Service, Intermountain Research Station: 558-571.
Stevens, R. 2004a. Incorporating wildlife habitat needs into restoration and rehabilitation projects. In: Monsen, S.B.; Stevens, R.; Shaw, N.L., eds. Restoring western ranges and wildlands. Gen. Tech. Rep. RMRS-GTR-136-vol. 1. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station: 155-174.
Stevens, R. 2004b. Management of restored and revegetated sites. In: Monsen, S.B.; Stevens, R.; Shaw, N.L., comps. Restoring western ranges and wildlands, vol. 1. Gen. Tech. Rep. RMRS-GTR-136-vol-1. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station: 193-198.
Stevens, R.; Jorgensen, K.R. 1994. Rangeland species germination through 25 and up to 40 years of warehouse storage. In: Monsen, S.B.; Kitchen, S.G., comps. Proceedings- Ecology and management of annual rangelands; 1992 May 18-22; Boise, ID. Gen. Tech. Rep INT-GTR-313. Ogden, UT: U.S. Department of Agriculture, Forest Service, Intermountain Research Station: 257-265.
Stevens, R.; Meyer, S.E. 1990. Seed quality testing for range and wildland species. Rangelands. 12(6): 341-346.
Stevens, R.; Monsen, S.B. 2004. Forbs for seeding range and wildlife habitats. In: Monsen, S.B.; Stevens, R.; Shaw, N.L., comps. Restoring western ranges and wildlands, vol. 2. Gen. Tech. Rep. RMRS-GTR-136-vol-2. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station: 425-466.
Stevens, R.J.; Jorgensen, K.R.; Young, A.S.; Monsen, S.B. 1996. Forb and shrub seed production guide for Utah. AG 501. Logan, UT: Utah State University Extension. 52 p.
Swoboda, K.A. 2007. The pollination ecology of Hedysarum boreale Nutt. (Fabaceae) and evaluation of its pollinating bees for restoration seed production. Logan, UT: Utah State University. Thesis. 117 p.
Swoboda, K.A.; Cane, J.H. 2012. Breeding biology and incremental benefits of outcrossing for the restoration wildflower, Hedysarum boreale Nutt. (Fabaceae). Plant Species Biology. 27(2): 138-146.
Taylor, R.J. 1992. Sagebrush country: A wildflower sanctuary. Missoula, MT: Mountain Press Publishing Company. 211 p.
Tepedino, V.J.; Stackhouse, M. 1987. Bee visitors of sweetvetch, Hedysarum boreale boreale (Leguminosae), and their pollen-collecting activities. Great Basin Naturalist. 47(2): 314-318.
Tilley, D.; Ogle, D. 2011. Using historic bushel weights to estimate seed quality. Native Plants Journal. 12(2): 107-110.
Tilley, D.; Wolf, M.; St. John, L. 2022. Establishment and 10-year persistence of plant materials at Curlew National Grassland in southern Idaho. Native Plants Journal. 23(2): 178-192.
Treshow, M.; Harper, K. 1974. Longevity of perennial forbs and grasses. Oikos. 25(1): 93-96.
Tyser, R.W. 1992. Vegetation associated with two alien plant species in a fescue grassland in Glacier National Park, Montana. The Great Basin Naturalist. 52(2): 189-193.
Uppala, S.; Wu, B.M.; Temple, T.N. 2013. First report of Fusarium solani on Utah sweetvetch in the United States. Plant Disease. 97(3): 423.
USDA Forest Service [USDA FS] 1937. Range plant handbook. Washington, DC: U.S. Department of Agriculture, Forest Service. 816 p.
USDA Forest Service, Bend Seed Extractory [USDA FS BSE]. 2025. Nursery Management Information System Version 4.1.11. Local Source Report 34-Source Received. Bend, OR: U.S. Department of Agriculture, Forest Service Bend Seed Extractory.
USDA Forest Service, Western Wildland Environmental Threat Assessment Center [USFS WWETAC]. 2024. TRM Seed Zone Applications. Prineville, OR: U.S. Department of Agriculture, Forest Service, Western Wildland Environmental Threat Assessment Center. Available: https://www.fs.usda.gov/wwetac/threat-map/TRMSeedZoneMapper.php
USDA Natural Resources Conservation Service [USDA NRCS]. 2025. The PLANTS Database. Greensboro, NC: U.S. Department of Agriculture, Natural Resources Conservation Service, National Plant Data Team. https://plants.usda.gov/home
USDI Bureau of Land Management, Seeds of Success [USDI BLM SOS]. 2023. Bureau of Land Management technical protocol for the collection, study, and conservation of seeds from native plant species for Seeds of Success. Washington, DC: U.S. Department of the Interior, Bureau of Land Management, Plant Conservation and Restoration Program. 45 p.
USDI Bureau of Land Management, Seeds of Success [USDI BLM SOS]. 2025. Seeds of Success collection data. Washington, DC: U.S. Department of the Interior, Bureau of Land Management, Plant Conservation and Restoration Program.
USDI Environmental Protection Agency [USDI EPA]. 2018. Ecoregions. Washington, DC: U.S. Environmental Protection Agency. https://www.epa.gov/eco-research/ecoregions
USDI Geological Survey [USDI USGS]. 2023. Biodiversity Information Serving Our Nation (BISON). U.S. Geological Survey. https://www.gbif.us/#home
Utah Crop Improvement Association [UCIA]. 2015. How to be a seed connoisseur. Logan, UT: UCIA, Utah Department of Agriculture and Food, Utah State University and Utah State Seed Laboratory. 16 p.
Utah State University. 2024. EcoRestore Utah. Logan, UT: Utah State University Extension. https://extension.usu.edu/ecorestore/
Walker, S.C.; Shaw, N.L. 2005. Current and potential use of broadleaf herbs for reestablishing native communities. In: Shaw, N.L.; Pellant, M.; Monsen, S.B., comps. Sage-grouse habitat restoration symposium proceedings; 2001 June 4-7; Boise, ID. RMRS-P-38. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station: 56-61.
Wasser, C.H.; Shoemaker, J. 1982. Ecology and culture of selected species useful in revegetating disturbed lands in the West. FWS/OBS–82/56. Washington, DC: U.S. Fish and Wildlife Service, Office of Biological Services, Western Energy and Land Use Team. 347.
Weber, W.A. 1976. Rocky Mountain flora. Boulder, CO: Colorado Associated University Press. 479 p.
Welsh, S.L.; Atwood, N.D.; Goodrich, S.; Higgins, L.C., eds. 2015. A Utah Flora. Fifth Edition, revised. Provo, UT: Brigham Young University. 990 p.
Wiese, J.L.; Meadow, J.F.; Lapp, J.A. 2012. Seed weights for northern Rocky Mountain native plants with an emphasis on Glacier National Park. Native Plants Journal. 13(1): 39-49.
Winslow, S.R. 2002. Propagation protocol for production of seeds Hedysarum boreale Nutt. seeds. Native Plant Network. U.S. Department of Agriculture, Forest Service, National Center for Reforestation, Nurseries, and Genetic Resources. http://npn.rngr.net/propagation/protocols [Accessed 2024 December 17].
Wojcik, V.; Morandin, L. 2008a. Selecting plants for pollinators: A guide for gardeners, farmers, and land managers in the Black Hills coniferous forest ecoregion. San Francisco, CA: Pollinator Partnership. 27 p. Available: https://www.pollinator.org/pollinator.org/assets/generalFiles/BlackHills.May.2024.rx7.pdf
Wojcik, V.; Morandin, L. 2008b. Selecting plants for pollinators: A guide for gardeners, farmers, and land managers in the Great Plains Palouse Dry Steppe province. San Francisco, CA: Pollinator Partnership. 27 p. Available: https://www.pollinator.org/pollinator.org/assets/generalFiles/Great.Plains.Palouse.Dry.Steppe.May.2024.rx5.pdf
Young, S.A.; Schrumpf, B.; Bouck, M.; Moore, M. 2020. How the Association of Official Seed Certifying Agencies (AOSCA) tracks wildland sourced seed and other plant propagating materials. Logan, UT: Utah State University, Utah Agricultural Experiment Station. 36 p.
How to Cite
Gucker, Corey L.; Shaw, Nancy L. 2025. Utah sweetvetch (Hedysarum boreale). In: Gucker, C.L.; Shaw, N.L., eds. Western forbs: Biology, ecology, and use in restoration. Reno, NV: Great Basin Fire Science Exchange. https://westernforbs.org