Authorship
Gucker, Corey; Shaw, Nancy
Publication Date
November 2024
Nomenclature
Sanddune wallflower (Erysimum capitatum [Douglas ex Hook] Greene) belongs to the Erysimeae tribe of the Brassicaceae or mustard family (Al-Shehbaz 2010).
Family
Brassicaceae – Mustard family
Genus
Erysimum
Species
capitatum
NRCS Plant Code
ERCA14 (USDA NRCS 2024).
Subtaxa
The Flora of North America recognizes two varieties of sanddune wallflower, E. c. var. capitatum and E. c. var. purshii (Al-Shehbaz 2010, 2020).
Synonyms
Sanddune wallflower is a highly variable species, and hybridization within the Erysimum genus is common, thus there are many synonyms (Table 1 and 2, Al-Shehbaz 2010).
Table 1. Synonym list for variety capitatum.
| Genus | Species (and variety) | Authority |
| Cheiranthus | angustatus | Greene |
| arkansanus | (Nuttall) Greene | |
| californicus | (Greene) Greene | |
| elatus | (Nuttall) Greene | |
| pacificus | E. Sheldon | |
| wheeleri | (Rothrock) Greene | |
| Cheirinia | arkansana | (Nuttall) Moldenke |
| elata | (Nuttall) Rydberg | |
| wheeleri | (Rothrock) Rydberg | |
| Erysimum | arkansanum | Nuttall |
| asperum var. arkansanum | (Nuttall) A. Gray | |
| asperum var. bealianum | Jepson | |
| asperum var. capitatum | (Douglas ex Hooker) B. Boivin | |
| asperum var. elatum | (Nuttall) Torrey | |
| asperum var. stellatum | J. T. Howell | |
| californicum | Greene | |
| capitatum var. angustatum | (Greene) Rossbach | |
| capitatum var. bealianum | (Jepson) Rossbach | |
| capitatum var. stellatum | (J. T. Howell) Twisselmann | |
| capitatum var. washoense | Rossbach | |
| elatum | Nuttall | |
| insulare var. angustatum | (Greene) Jepson | |
| moniliforme | Eastwood | |
| suffrutescens var. lompocense | Rossbach | |
| tilimii | J. Gay | |
| wheeleri | Rothrock |
Table 2. Synonym list for variety purshii (Al-Shehbaz 2010).
| Genus | Species (and variety) | Authority |
| Cheiranthus | alpestris | (Cockerell) A. Heller |
| argillosus | Greene | |
| aridus | A. Nelson | |
| asperrimus | Greene | |
| bakeri | Greene | |
| nivalis | Greene | |
| nivalis var. amoenus | Greene | |
| oblanceolatus | (Rydberg) A. Heller | |
| radicatus | (Greene) | |
| Cheirinia | amoena | (Greene) Rydberg |
| argillosa | (Greene) Rydberg | |
| arida | (A. Nelson) Rydberg | |
| asperrima | (Greene) Rydberg | |
| bakeri | (Greene) Rydberg | |
| brachycarpa | Rydberg | |
| cockerelliana | (Daniels) Cockerell | |
| desertorum | Wooton & Standley | |
| nivalis | (Greene) Rydberg | |
| nivalis var. radicata | (Rydberg) Cockerell | |
| oblanceolata | (Rydberg) Rydberg | |
| radicata | (Rydberg) Rydberg | |
| Erysimum | amoenum | (Greene) Rydberg |
| angustatum | Rydberg | |
| argillosum | (Greene) Rydberg | |
| aridum | (A. Nelson) A. Nelson | |
| asperrimum | (Greene) Rydberg | |
| asperum var. amoenum | (Greene) Reveal | |
| asperum var. angustatum | (Rydberg) B. Boivin | |
| asperum var. pumilum | (Greene) Rydberg | |
| bakeri | Rydberg | |
| capitatum var. amoenum | (Greene) R. J. Davis | |
| capitatum var. argillosum | (Greene) R. J. Davis | |
| capitatum var. nivale | (Greene) N. H. Holmgren | |
| cockerellianum | Daniels | |
| desertorum | (Wooton & Standley) Rossbach | |
| nivale | (Greene) Rydberg | |
| oblanceolatum | Rydberg | |
| radicatum | Rydberg |
Common Names
Sanddune wallflower, California wallflower, prairie rocket, western wallflower (Hermann 1966; Arnberger 1982; Hickman 1993; LBJWC 2023; Discover Life 2024).
Chromosome Number
2n = 36 (Al-Shehbaz 2010).
Hybridization
Almost all native Erysimum species hybridize in areas of overlap, which results in a multitude of intermediates and backcrossing that blur species boundaries (Al-Shehbaz 2010).
Price (1986) found 7.5 to 63% seed set in artificial crosses of sanddune wallflower with western wallflower (E. asperum), pale wallflower (E. occidentale), Santa Cruz wallflower (E. teretifolium), San Francisco wallflower (E. franciscanum), Menzies’ wallflower (E. menziesii), and curly wallflower (E. menziesii subsp. concinnum), but these species typically occupy different habitats in areas where their ranges overlap. For example, sanddune wallflower and pale wallflower populations occur within 0.6 to 1.2 mi (1-2 km) of one another in the Columbia River Gorge and the Oregon-Washington border, sanddune wallflower grows on basaltic cliffs and hillsides while pale wallflower grows along rivers. As such, the two species typically do not hybridize in these areas. Sanddune wallflower and western wallflower populations occur near one another along the Rocky Mountain Front Range from Montana to New Mexico, but the species remain distinct and separated ecologically and geographically. However, intermediate forms from hybridization between sanddune wallflower and island wallflower (E. insulare subsp. suffrutescens) occur on hillsides near coastal dunes in Ventura, Santa Barbara, and San Luis Obispo Counties in California (Price 1986).
Distribution
Sanddune wallflower is very widespread in terms of geographic area and habitat, with populations throughout the western half of the United States and as far east as Ohio and Tennessee (Table 3; Price 1986; Al-Shehbaz 2010). Populations can be widely scattered and disjunct. Of the two sanddune wallflower varieties, capitatum occupies the broadest range but it is only sporadically collected outside of its main range from the Pacific states through western Nevada and western Idaho. Variety purshii occurs in Alaska and Yukon and then in disjunct populations from Idaho and Montana south to California and westernmost Texas. It does not occur in Washington or Oregon (Al-Shehbaz 2010).
Table 3. Distribution of sanddune wallfower varieties by state or territory from Canada to Mexico (Price 1986; Al-Shehbaz 2010).
| State/Territory/Region | Variety |
| Alaska | purshii |
| Arizona | capitatum, purshii |
| Arkansas | capitatum |
| California | capitatum, purshii |
| Colorado | purshii |
| Idaho | capitatum, purshii |
| Illinois | capitatum |
| Indiana | capitatum |
| Iowa | capitatum |
| Montana | capitatum, purshii |
| Missouri | capitatum |
| New Mexico | purshii |
| Nevada | capitatum, purshii |
| Ohio | capitatum |
| Oklahoma | capitatum |
| Oregon | capitatum |
| Tennessee | capitatum |
| Texas | capitatum, purshii |
| Utah | purshii |
| Washington | capitatum |
| Wyoming | purshii |
| Yukon, CA | purshii |
| Chihuahua, MX | purshii |
| Durango, MX | capitatum |
| Michoacan, MX | purshii |
| San Luis Potosi, MX | capitatum |
| Sonora, MX | purshii |
| Zacatecas, MX | purshii |
Habitat And Plant Associations
Sanddune wallflower occupies sites from valley bottoms to alpine mountaintops. It occurs in desert grasslands (Fig. 1), desert shrub (Fig. 2), chaparral, aspen (Populus tremuloides), pinyon-juniper (Pinus–Juniperus spp.) woodlands (Fig. 3), forest openings, and alpine habitats (Price 1986; Blackwell 2006; Al-Shehbaz 2010; Lesica 2012; Spellenberg 2012).

Figure 1. Sanddune wallflower growing in a grassland habitat in California. Photo: Bureau of Land Management/University of California Botanical Garden (BLM/UCBG), Seeds of Success (SOS).

Figure 2. Sanddune wallflower growing in a Wyoming big sagebrush (Artemisia tridentata subsp. wyomingensis) habitat in Butte County, ID. Photo: Matt Lavin.

Figure 3. Sanddune wallflower growing in a mature juniper woodland in Utah. Photo: BLM UT060, SOS.
Because of sanddune wallflower’s low fidelity to plant associations, a small sample of its habitats reported in the literature is described here with more plant associations described in the soils section below. In the western Mojave, sanddune wallflower occurred on desert plains in the Joshua tree (Yucca brevifolia) zone, on rough deposits of more or less active hot springs in the upper Sororan Zone of Washoe County, Nevada, and where vegetation was sparse on sand dunes along the tidal estuary of San Joaquin River in Contra Costa County, California (Rossbach 1958). Sanddune wallflower was common on the Sanup Plateau (4,840 ft [1,475 m]) on the Grand Canyon’s northern rim where blackbrush (Coleogyne ramosissima), Mexican cliffrose (Purshia mexicana), and Utah agave (Agave utahensis) dominated (Phillips 1975). At high-elevation sites (12,300 ft [3,750 m]) on the East Slope of the Front Range in Boulder County, Colorado, sanddune wallflower grew in Bellardi bog sedge (Kobresia myosuroides) meadows experiencing high winds and short growing seasons (Kooiman and Linhart 1986). Near its northernmost range, sanddune wallflower occurred in grasslands of the Aishihik-Sekulmun Lakes area in Canada’s Yukon Territory. These grasslands occupied dry, steep, south-facing slopes dominated by prairie sagewort (Artemisia frigida) and threadleaf sedge (Carex filifolia) (Vetter 2000).
Elevation
Sanddune wallflower grows at elevations from sea level to 13,120 ft (4,000 m) (Hickman 1993; Al-Shehbaz 2010; Spellenberg 2012). Variety capitatum occurs at elevations from 0 to 5,600 ft (1,700 m) and variety purshii at elevations from 3,280 to 13,120 ft (1,000-4,000 m) (Al-Shehbaz 2010).
Soils
Sanddune wallflower often occurs on dry, sandy and gravelly or stony soils (Fig. 4) (Craighead et al. 1963; Munz and Keck 1973; Taylor 1992; Blackwell 2006; Lesica 2012; LBJWC 2023). Plants grow on many substrates including quartzite, quartz sand, argillite, quartz diorite, sandstone, granite, basalt, limestone, serpentine, and occasionally travertine, gypsum, or pumice (Whittaker 1960; Price 1986; Grace et al. 2007).

Figure 4. Sanddune wallflower with orange flowers growing on a very rocky site in California’s Sacramento Mountains. Photo: Wynn Anderson, Lady Bird Johnson Wildflower Center (LBJWC).
Several studies report sanddune wallflower growing in dry habitats. It was described as a xerophytic species on porous desert plains soils in the western Mojave Desert. It grew on more or less consolidated sand dunes of fine sand and some clay dust along the San Joaquin River in Contra Costa County, California (Rossbach 1958). It also occurred in loose sand of stabilized dunes and sandhills at elevations of 30 to 820 ft (9-250 m) in Santa Barbara County, California (Baldwin 2022). In a survey of 263 plots from montane to alpine zones in the Bishop Creek Watershed on the eastern side of the Sierra Nevada Mountains in California, sanddune wallflower was found in 19 plots that were significantly associated with dry conditions (P < 0.01) (Kimball et al. 2004).
In addition to various substrates, low organic matter and low fertility are also tolerated. In the Siskiyou Mountains of Oregon and California, sanddune wallflower was found from low to high elevations on xeric serpentine, quartzite, argillite, and quartz diorite soils (Whittaker 1960). In a study of vegetation-substrate relationships in the bristlecone pine (Pinus aristata) zone (9,500-11,000 ft [2,900-3,350 m]) of California’s White Mountains, sanddune wallflower occurred on granite but not on dolomite or sandstone soils in the plots sampled. Percent sand was greatest and percent soil moisture was lowest in granite soils in the cold, dry study area (Wright and Mooney 1965). In a study of topography, soils, and vegetation in northern Arizona, sanddune wallflower occurred on dry, sandy loam, red cinder soils low in organic carbon and total nitrogen (Abella and Covington 2006). Sanddune wallflower occurred in dry forests, dry meadows, and colluvium associations in Emerald Lake Basin in Sequoia National Park, California (Rundel et al. 2009). The basin was a glacial cirque carved from granitic material. Dry forests and meadows occupied shallow decomposed granite layers low in organic matter that dried early in the growing season. The colluvium association occupied steep slopes with shallow soils that developed between granitic boulders (Rundel et al. 2009). During a study of Charleston Mountain pussytoes (Antennaria soliceps), a narrow endemic on limestone talus, above and below the tree line in Nevada’s Spring Mountains, sanddune wallflower occurred on steep slopes with soils high in manganese (Bayer and Minish 1993).
Description
Sanddune wallflower is a highly variable species. Field identification can be difficult because most Erysimum species shed their basal leaves and lack flowers when distinguishing fruits are produced. Nearly all native Erysimum species can hybridize where distributions overlap, resulting in many intermediate forms and backcrosses (Al-Shehbaz 2010).
Sanddune wallflower plants grow as biennials or short-lived perennials throughout the species range (Al-Shehbaz 2010) and possibly as winter annuals in the Mojave Desert and southern California (Price 1986). Plants produce one to several erect stems that are often branched distally and sometimes proximally (Mansfield 2000; Blackwell 2006; Al-Shehbaz 2010; Spellenberg 2012). Stature, degree of branching, and life cycle vary by habitat (Figs. 5 and 6). Alpine plants are typically dwarf, multi-stemmed and branched, perennials. Plants growing at low-elevation sites are taller, less branched, and biennial (Price 1986). Plants are taprooted with the degree of lateral root development depending on firmness of the substrate (Price 1986; Lesica 2012). They develop short to elongate caudices that are often clothed in old leaf bases (Hickman 1993; Lesica 2012). Stems are coarse, leafy, and 0.5 to 4 ft (1.5-12 dm) tall (Munz and Keck 1973; Mansfield 2000; Blackwell 2006; Spellenberg 2012). Herbage is covered in stiff, appressed hairs with two or more forks or branches (Munz and Keck 1973; Hickman 1993). Plants leak a watery, pungent juice when tissue is damaged (Craighead et al. 1963).

Figure 5. Large sanddune wallflower plant growing in New Mexico. Photo: BLM NM018, SOS.

Figure 6. Dwarf sanddune wallflower growing in Wyoming. Photo: BLM WY030, SOS.
Plants produce simple basal and stem leaves with entire to dentate or denticulate margins (Fig. 7) (Al-Shehbaz 2010, 2020). Leaves are alternate (Taylor 1992). Basal leaves are spatulate to linear, 0.8 to 10.5 in (2-27 cm) long, and 0.1 to 1.2 in (3-30 mm) wide (Al-Shehbaz 2010, 2020). Stem leaves are sessile, angled strongly upward, and gradually reduced upwards (Lesica 2012; Spellenberg 2012). Leaf shape is variable, but all leaves have prominent midribs (Price 1986).

Figure 7. Sanddune wallflower with dentate leaf margins growing in California. Photo: BLM/UCBG, Seeds of Success.
Stems are topped with congested racemes (Munz and Keck 1973; Blackwell 2006; Spellenberg 2012). Individual flowers (1-1.5 in [2.5-3.8 cm] wide) are perfect with four separate round petals that form a cross (Blackwell 2006; Spellenberg 2012; Hitchcock and Cronquist 2018). Petals are usually yellow to orange (Figs. 4, 7) and occasionally lavender to maroon at high-elevation sites (Munz and Keck 1973; Arnberger 1982; Al-Shehbaz 2010, 2020). Flowers have stout styles up to 3 mm long and six stamens, four longer than the other two (Mansfield 2000). Racemes become much elongated in fruit (Lesica 2012). Fruits are linear, narrow siliques (1.4-6 in [3.5-15] cm long by 1.3-3.3 mm thick), mostly straight, and borne upright in the raceme (Al-Shehbaz 2010, 2020; Spellenberg 2012). Siliques are four-sided and slightly flattened with prominent midveins (Blackwell 2006; Spellenberg 2012). Pods are many seeded (40-80 ovules/valve) and split open when mature (Taylor 1992; Al-Shehbaz 2010, 2020; Hitchcock and Cronquist 2018). Individual seeds are oblong and about 1.5 mm long (Munz and Keck 1973; Al-Shehbaz 2010, 2020).

Figure 8. Sanddune wallflower inflorescence with developing siliques. Photo: Matt Lavin.
A study of four sites on the eastern slope of the Rocky Mountains in Colorado, shows how varied sanddune wallflower plants can be across a 3,000-ft (900 m) elevation range (Table 4a). Plants that produced taller inflorescences typically produced more and larger flowers than plants with shorter inflorescences (Lay et al. 2013).
Table 4a. Morphological characteristics (means) of sanddune wallflower across an elevational gradient in the Rocky Mountains of Colorado (Lay et al. 2013).
| Average across 2 years by site |
Inflorescence height (in) | Stem diameter (mm) | Open flowers (no.) | Corolla width (mm) |
| Short-grass prairie (6,500 ft) | 7.3c | 2.3a | 9.6a | 15.1b |
| Open ponderosa pine (7,250 ft) | 9.5a | 2.1a | 6.9b | 15.8a |
| Open ponderosa pine (8,230 ft) | 6.4d | 1.8c | 5.4c | 14.2c |
| Subalpine meadow (9,500 ft) | 8.2b | 1.9b | 5.3c | 15.1b |
Numbers within each column followed by different letters are significantly different (P < 0.05)
Distinguishing varieties.
Variety capitatum produces mostly three-rayed but up to seven-rayed hairs on the upper sides of basal and lower stem leaves. Flower petals are usually orange (Fig. 4) but can be orange-yellow to yellow. Siliques have fewer than four-angles. Seeds are winged, at least distally, and measure 2 to 4 mm by 1 to 2 mm (Al-Shehbaz 2010).
Variety purshii produces mostly two- or three-rayed hairs on the upper sides of basal and lower stem leaves. Flower petals are yellow and sometimes, but rarely, lavender or purplish. Siliques are four-angled. Seeds are not winged and measure 1.5 to 2.4 mm by 0.7 to 1.2 mm (Al-Shehbaz 2010).
Reproduction
Although sanddune wallflower grows as a biennial or short-lived perennial, plants often flower in their first year (LBJWC 2023). Pollination is required for seed production (Lay et al. 2011). Potential pollinators include bees, flies, and pollen beetles (Lay et al. 2013). Silique length is strongly correlated to seed set. Fruits failing to elongate fully contain only a few seeds (Price 1986).
Extensive demography and life history studies of sanddune wallflower populations were conducted across a range of elevations in the Rocky Mountains of Colorado (Kim and Donohue 2001a, 2011b, 2013). These studies found that populations were typically semelparous (reproducing once/lifetime) in dry, low-elevation sites, iteroparous (>1 reproductive event/lifetime) at moist, high-elevation sites, and some populations exhibited intermediate parity (Table 5, Kim and Donohue 2011b). The study included six populations. High-elevation populations (H1, H2, H3) occurred in alpine tundra, and low-elevation populations occurred in sagebrush (Artemisia spp.) scrub (L1) and open ponderosa pine (Pinus ponderosa) forests (L2, L3). Density of sanddune wallflower, especially that of new seedlings, was lower at high-elevation than low-elevation sites (P < 0.1). High-elevation sites had a larger proportion of adult plants than low-elevation sites (P < 0.001), and survival at all life stages was higher in high than low habitats in 2005 (P < 0.1). Average survival of reproductive individuals at high sites was six to seven times higher than at low sites in 2005. Summer soil moisture was lower at low than high sites, and soil water availability was positively associated with a population’s post-reproductive survival (Kim and Donohue 2011b).
Table 5. Life history strategies for sanddune wallflower growing at low and high elevation sites in Colorado’s Rocky Mountains (Kim and Donohue 2011a, 2011b).
| Population* | Elevation (ft) | Plant density (no/ft2) in 2004 | Survival rate of reproducing plants (%) | Life history |
| H1 | 10,470 | 0.4 | 87 | Iteroparous |
| H2 | 11,930 | 1.3 | 72 | Iteroparous |
| H3 | 11,500 | 1.7 | 20 | Intermediate |
| L1 | 8,630 | 1.7 | 17 | Intermediate |
| L2 | 7,330 | 1.6 | 0 | Semelparous |
| L3 | 6,000 | 1.8 | 3.5 | Semelparous |
*H1, H2, and H3 are populations from high-elevation, alpine tundra sites. L sites are populations from low-elevation sites in sagebrush (Artemisia spp.) scrub (L1) and open ponderosa pine (Pinus ponderosa) forests (L2, L3).
In follow-up studies, seeds collected from the same six populations were grown together in a greenhouse where water availability was varied (Kim and Donohue 2011a, 2012). Traits associated with iteroparity showed population differentiation and phenotypic plasticity in response to water availability. There were positive and negative effects of soil water content on post-reproductive survival, which could foster stable reproductive survival across variable environments where high populations could become more semelparous in dry conditions (Kim and Donohue 2011a). Nearly all plants (98.5%) survived the non-drought treatment and 34% survived the 6-month, drought-stressed treatment. Plants from low populations survived the drought stress conditions better than high populations (P < 0.1), which was attributed to differences in rosette size and number. Plants with fewer and smaller rosettes survived drought stress better (Kim and Donohue 2012).
Phenology
Depending on location, flowering of sanddune wallflower plants can occur from January to September, but it is most common from April to August (Munz and Keck 1973; Blackwell 2006; Al-Shehbaz 2010).
Pollination
Seed production requires pollination (Lay et al. 2011), and many insects visit sanddune wallflower flowers (Macior 1970; Alarcón et al. 2008; Lay et al. 2013). Plants produce an abundance of pollen grains and seeds based on estimates from populations growing in northern California, where 258,000 grains/anther, 38.3 ovules/flower, and pollen-ovule ratios of 6,800 were reported (Preston 1986).
Sanddune wallflower attracts a variety of pollinators. Flowers were commonly pollinated by flies, bees, and pollen beetles (Meligethes spp.) from short-grass prairie to the subalpine zone in the Front Range of Colorado (Lay et al. 2013). Flowers were repeatedly visited by western tiger swallowtail butterflies (Papilio rutulus) in a montane meadow in California’s San Bernadino Mountains (Alarcón et al. 2008). At a montane-alpine ecotone on the Front Range from Boulder to Park Counties in Colorado, sanddune wallflower was regularly pollinated by golden-belted bumble bees (Bombus balteatus) and forest bumble bees (B. sylvicola) (Macior 1970). See the Wildlife and Livestock Use section for a list of more pollinators and insect visitors.
Pollination was evaluated for sanddune wallflower populations at four sites on the eastern slope of the Rocky Mountains’ Front Range in 2005 and 2006 (Lay et al. 2013). Flowers were pollinated by flies, bees, and pollen beetles from 11 insect families. Flies were more common at higher elevation sites and bees and pollen beetles at lower elevations. A total of 244 total pollinators visited sanddune wallflower in the short-grass prairie (9,780 ft [2,980 m]) in 2040 minutes of observations, 222 pollinators in 2209 minutes and 88 in 1260 minutes of observations in ponderosa pine forests (7,250-8,240 ft [2,210-2,510 m]), and 244 pollinators in 2230 minutes of observations in dry subalpine meadows (9,500 ft [2,900 m]) (Lay et al. 2013).
Ecology
Sanddune wallflower is an early-seral species often growing on disturbed open areas, washes, and roadsides (Lesica 2012; Spellenberg 2012). Several studies report greater abundance of sanddune wallflower on grazed or burned than on undisturbed sites (Pearson 1965; Vogl and Schorr 1972).
Seed And Seedling Ecology
Sanddune wallflower seeds are small, lightweight (< 2mg each), and lack a mechanism for dispersal or self-burial (Pavlik and Manning 1993).
In-depth seed production and seed bank studies were conducted at Antioch Dunes National Wildlife Refuge in central California (Pavlik et al. 1993). In spring 1987, there were 2,207 sanddune wallflower plants, and in spring 1988, there were 845 sanddune wallflower plants at this site. This difference was attributed to poor germination and seedling establishment following the dry winter of 1986-87. In 1987 the maximum number of seeds produced by a single plant was 7,481, and the average per plant was 2,800. In 1988, the maximum number of seeds produced by a single plant was 383, and the average per plant was 192. In 1987 more than 80% of flowers produced fruits containing seed. The fruit/flower and seed/ovule ratios were reduced by 50% or more from 1987 to 1988 (Table 6, Pavlik et al. 1993).
Table 6. Fruit and seed production of sanddune wallflower* in a wetter (1987) and drier (1988) year at Antioch Dunes National Wildlife Refuge in central California (Pavlik et al. 1993).
| Study year | Fruit to flower ratio | Seed to ovule ratio |
| 1987 | 0.834 ± 0.102 b | 0.495 ± 0.061 b |
| 1988 | 0.417 ± 0.229 a | 0.125 ± 0.068 a |
Means within a column followed by different letters are significantly different (P < 0.01).
*Sanddune wallflower at this site was previously thought to be an endangered species but is now considered variety capitatum.
At Antioch Dunes, sanddune wallflower was visited and pollinated by a wide variety of insects (Pavlik et al. 1993). Pollinators were abundant over the study period, but the reduced number of plants in 1988 produced smaller inflorescences. Another factor limiting seed production was insect predation. Beetle (not identified) and moth (Chalceopla simpliciella) larvae reduced seed production by 13% in 1987 and 33% in 1988. In field experiments adding water and water plus nutrients to select plants in 1987, seed production was not increased, but these treatments were not tested in 1988 (Pavlik et al. 1993).
Seed bank.
The sanddune wallflower seed bank was also evaluated at Antioch Dunes (Table 7, Pavlik et al. 1993). About 20% of seeds remained in the residual seedbank in soil samples collected in December and May. Soil samples (28.1 in2 [181.5 cm2] surface area) were taken between mature reproductive plants (two samples) and beneath mature plants (three samples). Soil was collected in August 1987 just after developed fruits had begun to dehisce and release seeds, in December 1987 after heavy rains encouraged germination, in May 1988 before current year fruits had opened, and again in September 1988 in the seed dispersal period. Seed bank density was determined using the emergence method. At the study site, great brome (Bromus diandrus)-dominated seed bank emergence, outnumbering sanddune wallflower 3 to 1. Seed germination density of both was greatest in the fall (Pavlik et al. 1993). For information on an experimental seeding conducted at Antioch Dunes and an adjacent site, see Wildland Seed Certification section.
Table 7. Density of sanddune wallflower seed in soil collected over about 1 year at Antioch Dunes National Wildlife Refuge in central California (Pavlik et al. 1993).
| Collection date | Seed density (no/ft2) |
| August 1987 | 96 ± 90 |
| Dec 1987 | 21 ± 15 |
| May 1988 | 16 ± 11 |
| Sept 1988 | 67 ± 59 |
Disturbance Ecology
Studies indicate that sanddune wallflower is an early colonizer of disturbed sites (Fig. 9) (Stevens et al. 2019) and grows on newly formed soils (Rundel et al. 2009) but is also found later in succession on moving sand dunes. In a study of sand dune succession in Fremont County, Idaho, sanddune wallflower was noted on 80-year-old sand deposits in bitterbrush-rabbitbrush (Purshia–Chrysothamnus spp.) and on 133- to 500-year-old deposits in bitterbrush-sagebrush communities (Chadwick and Dalke 1965).
Herbivory.
Predation of Erysimum plants by insects and vertebrates is likely limited by the presence of cardenolides and glucosinolates. Yet, deer feed on young inflorescences, and Ceutorhynchus weevil larvae specialize on ripening seeds and have been found on up to 50% of the plants in some populations (Price 1986). When grazed and protected sites were compared in big sagebrush-needle-and-thread (Artemisia tridentata–Hesperostipa comata subsp. comata) habitats in Rexburg, Idaho, frequency of sanddune wallflower was 18% on sites grazed heavily for about 70 years and 4% on sites protected from grazing for 11 years (Pearson 1965).
The effects of herbivores on plant fitness were evaluated in 2005 and 2006 in four populations growing in short-grass prairie, open ponderosa pine forests, and subalpine meadows in Colorado’s Front Range. Sanddune wallflower plants that produced more flowers over the season and more open flowers per day had more pollinators and more herbivores (ants, gall midges, and petal consumers). Herbivory reduced pollinator visits in some but not all populations, but differences in pollinator visitation were not related to variations in reproductive success or fitness at any site or in either year. Pollen was not limited for sanddune wallflower suggesting that reproductive success (measured as silique length) was primarily limited by resource availability. Observations suggested that herbivory was a selective pressure. Stem borers preferred thicker stems, and at the site where stem borers were most common, sanddune wallflower plants had the narrowest stems of any population. At each population and in both years, researchers documented pollinator and herbivore visitation in relation to plant traits and effects of pollinator and herbivore visitation on plant fitness. Results of this tracking revealed compensatory responses by sanddune wallflower, which included replacement of damaged and unpollinated flowers and allocation to individual fruits (Lay et al. 2011).
Fire.
Sanddune wallflower is an early colonizer of burned sites (Fig. 9) (Stevens et al. 2019) and does not appear to be negatively affected by burning (Vogl and Schorr 1972; Fornwalt and Kaufmann 2014).

Figure 9. Robust sanddune wallflower plant growing and producing flowers in a 3-year-old burned Douglas-fir forest. Photo: Matt Lavin.
Sanddune wallflower colonized burned plots in the first postfire year following the Hayman fire in ponderosa pine-Douglas-fir (Pseudotsuga menziesii) forests on the Pike National Forest in Colorado on June 8, 2002. Plants occurred in the first postfire year and for 10 years after the fire on one moderate-severity burned and two low-severity burned plots. Low-severity burns saw less than 50% of overstory trees killed, and moderate-severity burns saw more than 50% of overstory trees killed (Stevens et al. 2019). In another evaluation of the Hayman fire, sanddune wallflower was consistently found in similar abundance on prefire and 1- to 5-year-old postfire plots (Fornwalt and Kaufmann 2014).
In Eastwood’s manzanita (Arctostaphylos glandulosa) vegetation in the San Jacinto Mountains south of Banning, California, sanddune wallflower occurred on burned but not unburned plots. The burned area was visited in the second postfire year following a late October fire. The researchers suggested all species found only on burned sites germinated from seed stored in the litter or soil (Vogl and Schorr 1972).
Wildlife And Livestock Use
Sanddune wallflower is eaten by a variety of large and small mammals and provides food and habitat for many insect species. Studies suggest that plants are occasionally grazed by elk (Cervus canadensis), bighorn sheep (Ovis canadensis) (Craighead et al. 1963), deer (Odocoileus spp.) ( McCulloch 1973; Price 1986), and domestic livestock (Herbel and Nelson 1966; Hermann 1966). In the analysis of deer taken in late winter from the Three Bar Wildlife Area in the Tonto National Forest, Arizona, sanddune wallflower occurred in 14% of white-tailed deer (O. virginianus) stomachs. The greatest volume was 4% in one sample (McCulloch 1973). On the Jornada Experimental Range north of Las Cruces, New Mexico, sanddune wallflower was eaten in the fall by cattle in one of three years of observations (Herbel and Nelson 1966).
Small mammals that feed on the stems and leaves of sanddune wallflower include pikas (Ochotona spp.), marmots (Marmota spp.), and desert cottontails (Sylvilagus audubonii) (Craighead et al. 1963; Turkowski 1975). Stems and leaves occurred in 7% of the stomachs of desert cottontails collected over a 15-month period at Sugarloaf Mountain in the Tonto National Forest, Arizona. At this yellow paloverde (Parkinsonia microphylla)-dominated site, the highest season preference index (average % volume per stomach/relative % ground cover) for desert cottontails was 11.66 from May to June 1972 (Turkowski 1975).
Sanddune wallflower is an important pollinator and host plant. The following insects are pollinator associates: miner bees (Andrena amphibola, A. cressonii, A. vicinoides), American bumble bee (Bombus pensylvanicus), sweat bees (Halictus tripartitus, Lasioglossum albipenne, L. cooleyi, L. perpunctatum, L. pruinosum, L. ruidosense, L. semicaeruleum, L. sisymbria, L. trizonatum, singular pebble bee (Dianthidium singular), leafcutter bees (Megachile angelarum), and mason bees (Osmia cordata parasitica) (Discover Life 2013).
Sanddune wallflower is used as a host plant by Sara orangetip (Anthocharis sara), large marble (Euchloe ausonides), and cabbage white (Pieris rapae) butterflies (Robinson 2024) and is a nectar source for adult Olympia marble butterflies (E. olympia) (Scott 2014 cited in Toliver 2022).
Sanddune wallflower was also identified as a key connector plant in bee-flower networks even though it was not one of the most abundant plants in recently thinned (3-10 yrs ago) ponderosa pine forests (6,890 to 9,190 ft [2,100-2,800 m]) in central Colorado’s Front Range. Connector plant species were those that supported modularity in bee-flower networks by linking interacting species together. Connector plants are recommended to receive high conservation priority due to their contributions to pollination network stability (Davies et al. 2023).
Nutritional Value
The sugar content of sanddune wallflower nectar was evaluated on 2-year-old burned sites on the Stanislaus National Forest, Tuolumne, California. From a sample of four flowers, sanddune wallflower nectar had an average of 0.007 mg of fructose, 0.004 mg of glucose, and 0.003 mg of sucrose for a total sugar mass of 0.014 mg (Pavlik et al. 2018).
Ethnobotany
Erysimum comes from the Greek word eryomai, which means to save. This background suggests medicinal properties (Mansfield 2000), and many indigenous groups describe medicinal uses of the plant. A poultice made by pounding the whole plant was applied to open, fresh wounds by the Okanagan-Colville (Turner et al. 1980; Moerman 2003). The Sioux made an infusion with crushed seed that was used externally for stomach or bowel cramps (Densmore 1918 cited in Moerman 2003).
Ramah Navaho used the plant as a ceremonial emetic, inhaled pulverized fruits pods to induce sneezing to relieve a congested nose, breathed in crushed leaves to treat headaches, applied heated roots to toothaches, and chewed the plant then exhaled the resulting fumes over a woman experiencing difficult labor (Vestal 1952). Zuni people ground the whole plant, mixed it with water, and applied the mixture to the forehead and temples to relieve pain from heat exhaustion and used it as a preventative for sunburn (Arnberger 1982). Flowers and fruits were used to induce vomiting to relieve stomach aches, and an infusion of the whole plant was applied topically for muscle aches by the Zuni (Camazine and Bye 1980). Hopi people used sanddune wallflower to treat advanced cases of tuberculosis (Colton 1974 cited in Moerman 2003). The Acoma and Laguna Pueblo Indians chewed leaves into a poultice for swellings and used the flowers to make yellow paint (Swank 1932).
Horticulture
Sanddune wallflower is commercially available for garden use. It is a showy plant that germinates readily (LBJWC 2023). It is recommended for USDA hardiness zones 3 to 9 and does well in sunny spots with well-drained soil (Missouri Botanical Garden 2024; PFAF 2024). Plants are bright colored, drought tolerant, attractive to pollinators, and easily grown from seed. Plants also have a long flowering period and their bright colored flowers look “particularly dazzling” against a dark backdrop (Stark 2020).
The following plant and care descriptions are offered by commercial seed sellers. Klamath-Siskiyou Native Seeds (2024) notes that sanddune wallflower does well in part shade or full sun in well-drained rocky to sandy soils and that removal of the top of the first bolting stem stimulates more flowering. Seeds can be sown directly outdoors from fall to early spring. Missouri Botanical Garden (2024) reports it is an easy-to-grow, low-maintenance plant in average dry to medium well-drained soils in full sun or light shade with good self-seeding when seedheads are allowed to mature. Sparrowhawk Native Plants (2024) reported that “nature turned up the color saturation filter up on [sanddune wallflower]”. It is a “joyful, pollinator-friendly option” and host and larval food source for several nocturnal moths. A short shelf life for purchased seed was noted by Stark (2020) with a recommendation to plant within 2 years of purchase.
Revegetation Use
As an early seral colonizer with good seed production and reestablishment potential and high pollinator value, sanddune wallflower has potential for use as a restoration species (see Pollination, Ecology, and Wildlife and Livestock Use). Its use as a revegetation species, however, has been extremely limited based on the reviewed literature.
Developing A Seed Supply
For restoration to be successful, the right seed needs to be planted in the right place at the right time. Coordinated planning and cooperation is required among all partners to select appropriate species and seed sources; these are determined by site conditions and current or projected revegetation requirements and goals (PCA 2015) and consultation with land managers and stakeholders.
Developing a seed supply begins with seed collection from native stands. Collected seed is distributed for restoration or production of nursery stock. Increase in agricultural seed fields may be required to provide a supply of seed (PCA 2015). Production of nursery stock requires less seed than large-scale seeding operations. Regardless of the size and complexity of any revegetation effort, seed certification is essential for tracking seed origin from collection through use (UCIA 2015).
Seed Sourcing
Researchers conducted a reciprocal planting field study between high-elevation populations (H1, H2, H3) in alpine tundra and low-elevation populations in sagebrush scrub (L1) and open ponderosa pine forests (L2, L3) at sites in the Colorado Front Range (Kim and Donohue 2013). The study included the use of seeds and seedlings from each population in pots placed in field locations. Seeds from H sites germinated better than seeds from L sites at all locations, but a lower proportion of their seeds germinated at L than H sites. L sites were associated with lower survival especially for H plants. Study findings overall found that sanddune wallflower exhibited home site advantage, survival was higher at native sites, especially for H plants, though some phenotypic plasticity in response to elevation was detected (Kim and Donohue 2013). See the reproduction section for more on this study.
Because empirical seed zones are not currently available for sanddune wallflower, generalized provisional seed zones developed by Bower et al. (2014) may be used to select and deploy seed sources. These provisional seed zones identify areas of climatic similarity with comparable winter minimum temperature and aridity (annual heat to moisture index). In Figure 10, Omernik Level III Ecoregions (Omernik 1987) overlay the provisional seed zones to identify climatically similar but ecologically different areas. For site-specific disturbance regimes and restoration objectives, seed collection locations within a seed zone and ecoregion may be further limited by elevation, soil type, or other factors.
The Western Wildland Environmental Threat Assessment Center’s (USDA FS WWETAC 2017) Threat and Resource Mapping (TRM) Seed Zone application provides links to interactive mapping features useful for seed collection and deployment planning. The Climate Smart Restoration Tool (Richardson et al. 2020) can also guide revegetation planning, seed collection, and seed deployment, particularly when addressing climate change considerations.
Occurrence Map
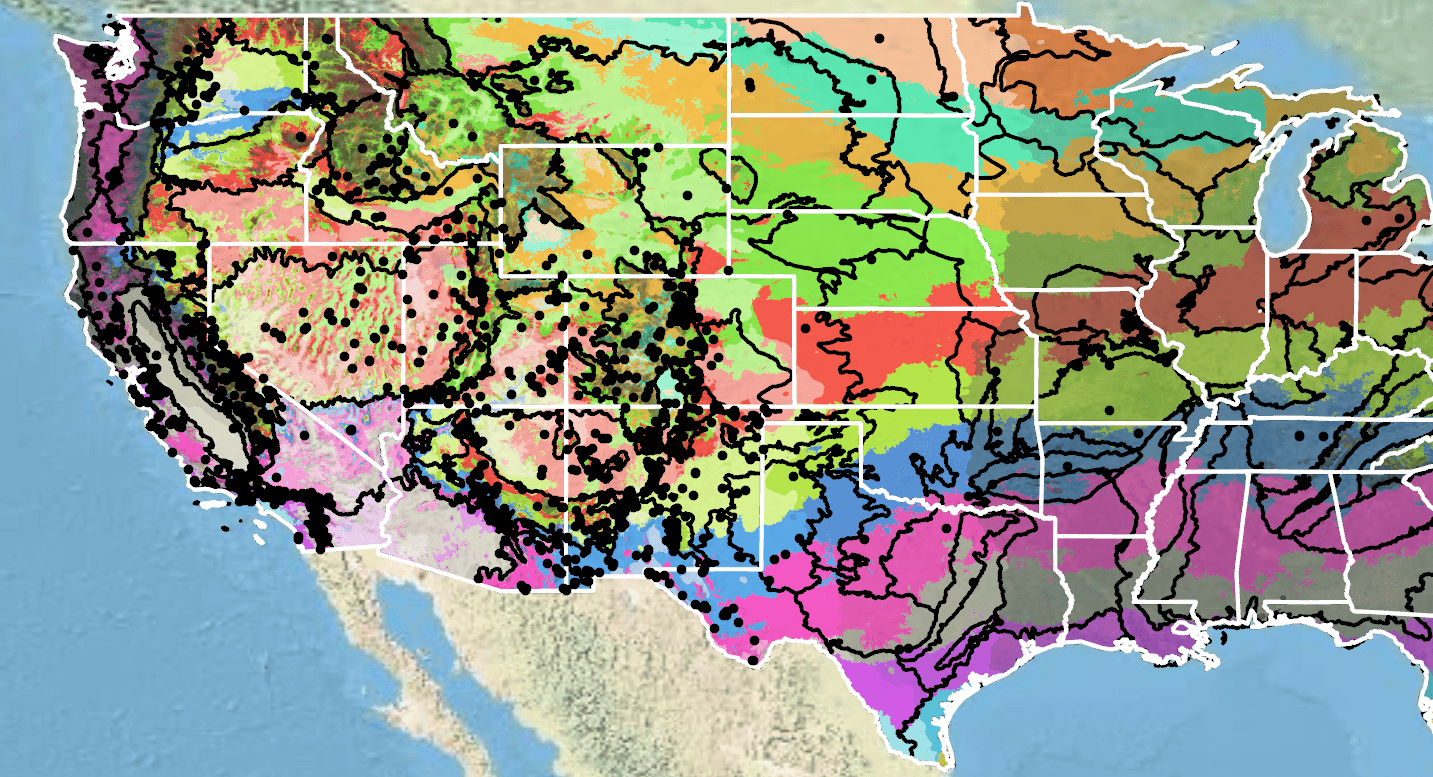
Figure 10. Distribution of sanddune wallflower in the contiguous United States (black circles) based on geo-referenced herbarium specimens and observational data from 1851–2019 (SEINet 2017; CPNWH 2020; USDI USGS 2020). Generalized provisional seed zones (colored regions) (Bower et al. 2014) are overlain by Omernik Level III Ecoregions (black outlines) (Omernik 1987; USDI EPA 2018). Interactive maps, legends, and a mobile app are available (USDA FS WWETAC 2017; research.fs.usda.gov/pnw/products/dataandtools/seed-zone-webmap). Map prepared by Sarah Barga and Benton Spendlove USFS RMRS.
Releases
As of 2024, there were no sanddune wallflower germplasm releases.
Wildland Seed Collection
Sanddune wallflower siliques open to release seeds when mature, making monitoring important to collecting any quantity of seed from this indeterminate species (Mansfield 2000). Although collection methods or tips were not reported in the reviewed literature, hand stripping is an effective method for collecting siliques of other species.

Figure 10. Sanddune wallflower seed stripped from of a single silique. Photo: BLM OR110, Seeds of Success.
Wildland Seed Certification
Verification of species and tracking of geographic source is necessary whether wildland seed is collected for immediate project use or as stock seed for cultivated increase. This official Source Identification process can be accomplished by following procedures established by the Association of Official Seed Certifying Agencies (AOSCA) Pre-Variety Germplasm Program (UCIA 2015; Young et al. 2020). Wildland seed collectors should become acquainted with state certification agency procedures, regulations, and deadlines in the states where they collect.
If wildland-collected seed is to be sold for direct use in ecological restoration projects, collectors must apply for Source-Identified certification prior to making collections. Pre-collection applications, site inspections, and species and seed amount verification are handled by the AOSCA member state agency where seed collections will be made (see listings at AOSCA.org).
If wildland seed collected by a grower or private collector is to be used as stock seed for planting cultivated seed fields or for nursery propagation (See Agricultural Seed Field Certification section), detailed information regarding the collection site and collecting procedures must be provided when applying for certification. Photos and herbarium specimens may be required. Germplasm accessions acquired within established protocols of recognized public agencies, however, are normally eligible to enter the certification process as stock seed without routine certification agency site inspections. For contract grow-outs, however, this collection site information must be provided to the grower to enable certification.
Collection Timing
Based on collection data from the BLM Seeds of Success collection crews, the harvest window is broad for sanddune wallflower (USDI BLM SOS 2024). The SOS made 9 collections in 2004, 2012, and from 2014 to 2016 in Colorado, Wyoming, Oregon, Utah, and New Mexico. The earliest collection was made on April 11, 2016, in Uintah County, Utah, at an elevation of 6,200 ft (1,890 m). The latest collection was made on August 25, 2014, from Platte County, Wyoming, at an elevation of 4,993 ft (1,522 m). Teams reported one collection in April, one in June, three in July and four in August. Collections spanned an elevation range of 1,360 ft (415 m) in Josephine County, Oregon when seed was harvested in mid-July 2012 to 9,743 ft (2,970 m) in Douglas County, Colorado when seed was collected in mid-August 2017 (USDI BLM SOS 2024).
Collection Methods
Although collection methods were not reported in the literature, it is likely that it is collected like that of other species producing silique-like fruits (see American vetch).
Several collection guidelines and methods should be followed to maximize the genetic diversity of wildland collections: 1) collect seed from a minimum of 50 randomly selected plants; 2) collect from widely separated individuals throughout a population without favoring the most robust or avoiding small stature plants; and 3) collect from all microsites including habitat edges (Basey et al. 2015). General collecting recommendations and guidelines are provided in online manuals (e.g., ENSCONET 2009; USDI BLM SOS 2023).
It is critical that wildland seed collection does not impact the sustainability of native plant populations. Collectors should take no more than 20% of the viable seed available at the time of harvest (USDI BLM SOS 2023). Additionally, care must be taken to avoid the inadvertent collection of weedy species, particularly those that produce seeds similar in shape and size to those of sanddune wallflower.
Post-Collection Management
Seed should be kept in a dry, shaded place until collections can be moved to a controlled short-term storage environment. Short-term storage should be dry, cool, and inaccessible to rodents or other seed predators. If insects are suspected in any collection, seed should be frozen for 48 hours or treated with an appropriate insecticide. The more plant material in the collection, the more ventilation and drying a seed lot will likely need (Gold n.d.; Parkinson and DeBolt 2005; Hay and Probert 2011).
Seed Cleaning
The following procedure was used to clean a 6.5-lb (2.9 kg) lot of sanddune wallflower seed (Fig. 11) grown in production plots at Lucky Peak Nursery, near Boise, Idaho (Barner 2009). Fruits were first processed through a Westrup Model LA-H laboratory brush machine (Hoffman Manufacturing, Corvallis, OR) with a #40 mantel and speed of 3 to remove seeds from the capsules. Seed was then air-screened to remove the remaining chaff and inert material using an office Clipper with a 1/14 or 1/16 top screen and 30 × 30 wire bottom screen, and medium speed and medium air. Personnel at the Bend Seed Extractory noted that farmed seed may be processed differently than native collections (Barner 2009).

Figure 11. Individual sanddune wallflower seeds about 2 mm long. Photo: BLM UT060, SOS.
Seed Storage
Sanddune wallflower seed is orthodox, and long-term storage (no length of time reported) was reported for ultra-dry seeds (no moisture content reported) stored at 14 °F (-10 °C) [SER, INSR, RBGK 2024]. The Bend Seed Extractory stored sanddune wallflower seed at 33 to 38 °F (0.5-3 °C) (Barner 2009).
Seed Testing
There are no AOSA purity or viability testing or germination guidelines for sanddune wallflower (AOSA 2010, 2023).
Germination Biology
The few germination studies for sanddune wallflower suggest that seed is nondormant and germinates with light and warm temperatures (Table 8, Maguire and Overland 1959).
Table 8. Germination percentages for sanddune wallflower seed collected in Washington and stored up to a year (Maguire and Overland 1959).
| Light, dark conditions | Dark | Dark-light | Dark | Dark | Dark-light |
| Temperature (°F) | 59 | 59 | 68 | 68-86 | 68-86 |
| Germination % | 36 | 56 | 52 | 48 | 62 |
In a greenhouse, sanddune wallflower germinated slightly better when watered with a 10% aqueous smoke solution (81%) than with just water alone (72%). Sanddune wallflower seed was collected from a ponderosa pine forest in Arizona, stored for 4 months at (23 °F [-5 °C]), and emergence was monitored after seed was planted in sterile potting soil kept in a greenhouse at 75 °F (24 °C) on a 14 hr/10 hr light-dark cycle. Emergence was 9% better for seed watered with a 10% aqueous smoke solution than for seed watered with water (Abella 2009).
Wildland Seed Yield And Quality
Post-cleaning seed yield and quality of seed lots collected in the Intermountain region are provided in Table 9 (USDA FS BSE 2021). The results indicate that sanddune wallflower seed can generally be cleaned to very high levels of purity and seed fill and that the viability of fresh seed is generally high.
Table 9. Seed yield and quality of sanddune wallflower seed lots collected in the Intermountain region, cleaned by the Bend Seed Extractory, and tested by the Oregon State Seed Laboratory or the USDA Forest Service National Seed Laboratory (USDA FS BSE 2021).
| Seed lot characteristic | Mean | Range | Samples (no.) |
| Bulk weight (lbs) | 0.39 | 0.08-1.27 | 13 |
| Clean weight (lbs) | 0.12 | 0.015-0.50 | 13 |
| Clean-out ratio | 0.30 | 0.06-0.95 | 13 |
| Purity (%) | 98 | 95-99 | 13 |
| Fill (%)¹ | 94 | 78-99 | 13 |
| Viability (%)² | 94 | 85-98 | 13 |
| Seeds/lb | 926,202 | 445,579-1,501,986 | 13 |
| Pure live seeds/lb | 846,733 | 379,366-1,334,250 | 13 |
¹ 100 seed X-ray test
² Tetrazolium chloride test
Marketing Standards
Acceptable seed purity, viability, and germination specifications vary with revegetation plans. Purity needs are highest for precision seeding equipment used in nurseries, while some rangeland seeding equipment handles less clean seed quite well.
Agricultural Seed Production
There were no reports of growing sanddune wallflower in an agricultural setting for seed increase.
Agricultural Seed Certification
In order to minimize genetic changes in specific accessions of native species when increased in cultivated fields, it is essential to track the geographic source and prevent inadvertent hybridization or selection pressure. This is accomplished by following third party seed certification protocols for Pre-Variety Germplasm (PVG) as established by the Association of Official Seed Certification Agencies (AOSCA). AOSCA members in the U.S., Canada, and other countries administer PVG requirements and standards that track the source and generation of planting stock. Field and cleaning facility inspections then monitor stand establishment, proper isolation distances, control of prohibited weeds, seed harvesting, cleaning, sampling, testing, and labeling for commercial sales (UCIA 2015; Young et al. 2020).
Seed growers apply for certification of their production fields prior to planting and plant only certified stock seed of an allowed generation (usually less than four). The systematic and sequential tracking through the certification process requires preplanning, knowing state regulations and deadlines, and is most smoothly navigated by working closely with state certification agency personnel. See the Wildland Seed Certification section for more information on stock seed sourcing.
Nursery Practice
Specific guidelines for growing sanddune wallflower container stock were not reported in the literature, but several studies utilized container plants in their studies (Abella 2009; Kim and Donohue 2011a, 2012) and none reported any difficulty.
Wildland Seeding And Planting
Based on the seeds/lb estimates in Table 9 above, the Natural Resources Conservation Service would recommend a seeding rate of 2.5 lb/ac (2.8 kg/ha) (to be adjusted by this species’ percentage of the total seeding mix) (D. Tilley, Personal Communication, September 2024).
An experimental field seeding project was conducted at sites adjacent to the Antioch Dunes National Wildlife Refuge in Contra Costa County, California (Table 9, Pavlik and Manning 1993). This area supports a sensitive sand dune ecosystem where sanddune wallflower grows. At the time of the study, the sanddune wallflower populations were considered an endangered subspecies, but they are now recognized as variety capitatum, a non-threatened or endangered plant. At the Refuge, sanddune wallflower grows on steep north-facing slopes with less sun exposure than south slopes. Population sizes fluctuated from 800 to 2,200 plants from 1985 to 1989. In November 1987, researchers established four experimental field sites: two on stabilized dunes with deep, sandy soils and two in adjacent clay soils disturbed by sand mining and vehicle compaction. The clay soils were mechanically disked to about 7.9 in (20 cm) deep. One of the dune sites was north facing with deep shade half of the day in fall, winter, and spring. The south-facing dune site was exposed and received little to no shading. Seed was sown in November 1987 at all four sites. Emergence occurred at all sites, but only at the dune sites did plants survive to reproductive age. More plants reached this stage at the shaded dune than at the exposed dune site. Researchers hypothesized that sanddune mortality was caused by water stress. The study area experienced drought conditions in 1988, which started in February and worsened with above average summer temperatures (>113 °F [45 °C]). Annual precipitation was 30 to 40% of normal during the study (Pavlik and Manning 1993).
Table 9. Emergence and survival of sanddune wallflower seeded at four sites with different exposure and substrates at Antioch Dunes National Wildlife Refuge in Contra Costa County, California (Pavlik and Manning 1993).
| Site | Seeds | Plants | ||||
| Number sown in Nov 1987 | Max germ (by 20 days after sowing) | Alive Feb 1988 | Alive May 1988 | Alive Sept 1988 | Reproductive May 1989 | |
| Dune (shaded) | 294 | 152a | 144a | 66a | 39a | 36a |
| Dune (exposed) | 294 | 140a | 113a | 54a | 3b | 1b |
| Clay (disked) | 294 | 109a | 105a | 74a | 0b | 0b |
| Clay (undisked) | 294 | 61b | 21b | 3b | 0b | 0b |
Numbers within a row for plant data followed by different letters are significantly different (P < 0.05).
Acknowledgements
Funding for Western Forbs: Biology, Ecology, and Use in Restoration was provided by the USDI BLM Great Basin Native Plant Materials Ecoregional Program through the Great Basin Fire Science Exchange. Great thanks to the chapter reviewers: Derek Tilley, Natural Resources Conservation Service, Aberdeen Plant Materials Center and Kim Frymire, U.S. Fish and Wildlife Service.
This research was supported in part by the USDA Forest Service, Rocky Mountain Research Station. The findings and conclusions in this publication are those of the authors and should not be construed to represent any official USDA or U.S. Government determination or policy.
Literature Cited
Abella, S.R. 2009. Smoke-cued emergence in plant species of ponderosa pine forests. Fire Ecology. 5(1): 22-37.
Abella, S.R.; Covington, W.W. 2006. Vegetation-environment relationships and ecological species groups of an Arizona Pinus ponderosa landscape, USA. Plant Ecology. 185(2): 255-268.
Al-Shehbaz, I.A. 2010. 378. Erysimum. In: Flora of North America Editorial Committee, ed. Flora of North America North of Mexico. Volume 7 Magnoliophyta: Salicaceae to Brassicaceae. New York, NY: Oxford University Press: 534-542.
Al-Shehbaz, I.A. 2020. Erysimum (Wallflower). In: Meyers, S.C.; Jaster, T.; Mitchell, K.E.; Harvey, T.; Hardison, L.K., eds. Flora of Oregon, Vol. 2: Dicots A-F. Fort Worth, TX: Botanical Research Institute of Texas (BRIT) Press: 483-486.
Alarcón, R.; Waser, N.M.; Ollerton, J. 2008. Year-to-year variation in the topology of a plant-pollinator interaction network. Oikos. 117(12): 1796-1807.
Arnberger, L.P. 1982. Flowers of the southwest mountains. Tucson, AZ: Southwest Parks and Monuments Association. 139 p.
Association of Official Seed Analysts [AOSA]. 2010. AOSA/SCST tetrazolium testing handbook. Contribution No. 29. Lincoln, NE: Association of Official Seed Analysts.
Association of Official Seed Analysts [AOSA]. 2023. AOSA rules for testing seeds. Vol. 1. Principles and procedures. Washington, DC: Association of Official Seed Analysts.
Baldwin, B.G. 2022. A new species of Layia (Compositae) from the central coast of California. Madrono. 69(1): 88-94.
Barner, J. 2009. Propagation protocol for production of propagules (seeds, cuttings, poles, etc.) Erysimum capitatum (Douglas ex Hook. [Greene]) seeds. Native Plant Network. U.S. Department of Agriculture, Forest Service, National Center for Reforestation, Nurseries, and Genetic Resources. http://npn.rngr.net/propagation/protocols [Accessed 2024 May 10].
Basey, A.C.; Fant, J.B.; Kramer, A.T. 2015. Producing native plant materials for restoration: 10 rules to collect and maintain genetic diversity. Native Plants Journal. 16(1): 37-53.
Bayer, R.J.; Minish, T.M. 1993. Isozyme variation, ecology, and phytogeography of Antennaria soliceps (Asteraceae: Inuleae), an alpine apomict from the Spring Mountains, Nevada. Madrono. 40(2): 75-89.
Blackwell, L.R. 2006. Great Basin wildflowers: A guide to common wildflowers of the high deserts of Nevada, Utah, and Oregon. Helena, MT: Morris Book Publishing. 288 p.
Bower, A.D.; St. Clair, J.B.; Erickson, V. 2014. Generalized provisional seed zones for native plants. Ecological Applications. 24(5): 913-919.
Camazine, S.; Bye, R.A. 1980. A study of the medical ethnobotany of the Zuni Indians of New Mexico. Journal of Ethnopharmacology. 2(4): 365-388.
Chadwick, H.W.; Dalke, P.D. 1965. Plant succession on dune sands in Fremont County, Idaho. Ecology. 46(6): 766-780.
Consortium of Pacific Northwest Herbaria [CPNWH]. 2020. Seattle, WA: University of Washington Herbarium, Burke Museum of Natural History and Culture. pnwherbaria.org/
Craighead, J.J.; Craighead, F.C. Jr.; Davis, R.J. 1963. A field guide to Rocky Mountain wildflowers from northern Arizona and New Mexico to British Columbia. Boston, MA: Houghton Mifflin Company. 277 p.
Davies, C.B.; Davis, T.S.; Griswold, T. 2023. Forest restoration treatments indirectly diversify pollination networks via floral- and temperature-mediated effects. Ecological Applications. 33(8): e2927
Discover Life. 2024. Sam Houston State University. Available: https://www.discoverlife.org/
European Native Seed Conservation Network [ENSCONET]. 2009. ENSCONET seed collecting manual for wild species. Edition 1: 32 p.
Fornwalt, P.J.; Kaufmann, M.R. 2014. Understorey plant community dynamics following a large, mixed severity wildfire in a Pinus ponderosa–Pseudotsuga menziesii forest, Colorado, USA. Journal of Vegetation Science. 25(3): 805-818.
Gold, K. n.d. Post-harvest handling of seed collections. Technical Information Sheet 04. UK: Royal Botanic Gardens Kew and Millennium Seed Bank Partnership. 4 p.
Grace, J.B.; Safford, H.D.; Harrison, S. 2007. Large-scale causes of variation in the serpentine vegetation of California. Plant Soil. 293: 121-132.
Hay F.R.; Probert, R.J. 2011. Chapter 20: Collecting and handling seeds in the field. In: Guarino, L.; Ramanatha, V.; Goldberg, E. Collecting plant genetic diversity: Technical Guidelines-2011 update. Rome, Italy: Bioversity International. 33 p.
Herbel, C.H.; Nelson, A.B. 1966. Species preference of Hereford and Santa Gertrudis cattle on a southern New Mexico range. Journal of Range Management. 19(4): 177-181.
Hermann, F. 1966. Notes on western range forbs: Cruciferae through Compositae. Agric. Handb. 293. Washington, DC: U.S. Department of Agriculture, Forest Service. 365 p.
Hickman, J.C., ed. 1993. The Jepson manual: Higher plants of California. Berkeley, CA: University of California Press. 1400 p.
Hitchcock, C.L.; Cronquist, A. 2018. Flora of the Pacific Northwest: An illustrated manual. Second Ed. Giblin, D.E.; Legler, B.S.; Zika, P.F.; Olmstead, R.G., eds. Seattle, WA: University of Washington Press. 882 p.
Kim, E.; Donohue, K. 2011a. Population differentiation and plasticity in vegetative ontogeny: Effects on life-history expression in Erysimum capitatum (Brassicaceae). American Journal of Botany. 98(11): 1752-1761.
Kim, E.; Donohue, K. 2011b. Demographic, developmental, and life-history variation across altitude in Erysimum capitatum. Journal of Ecology. 99(5): 1237-1249.
Kim, E.; Donohue, K. 2012. The effect of plant architecture on drought resistance: Implications for the evolution of semelparity in Erysimum capitatum. Functional Ecology. 26(1): 294-303.
Kim, E.; Donohue, K. 2013. Local adaptation and plasticity of Erysimum capitatum to altitude: Its implications for responses to climate change. Journal of Ecology. 101(3): 796-805.
Kimball, S.; Wilson, P.; Crowther, J. 2004. Local ecology and geographic ranges of plants in the Bishop Creek Watershed of the eastern Sierra Nevada, California, USA. Journal of Biogeography. 31(10): 1637-1657.
Klamath-Siskiyou Native Seeds. 2024. https://klamathsiskiyouseeds.com/ [Accessed 2024 July 17].
Kooiman, M.; Linhart, Y.B. 1986. Structure and change in herbaceous communities of four ecosystems in the Front Range, Colorado, U.S.A. Arctic and Alpine Research. 18(1): 97-110.
Lady Bird Johnson Wildflower Center [LBJWC]. 2023. Erysimum capitatum (Douglas ex Hook.) Greene. Native Plant Database. Austin, TX: Lady Bird Johnson Wildflower Center. https://www.wildflower.org/plants-main [Accessed 2024 July 17].
Lay, C.R.; Linhart, Y.B.; Diggle, P.K. 2011. The good, the bad and the flexible: Plant interactions with pollinators and herbivores over space and time are moderated by plant compensatory responses. Annals of Botany. 108(4): 749-763.
Lay, C.R.; Linhart, Y.B.; Diggle, P.K. 2013. Variation among four populations of Erysimum capitatum in phenotype, pollination and herbivory over an elevational gradient. The American Midland Naturalist. 169(2): 259-273.
Lesica, P. 2012. Manual of Montana vascular plants. Fort Worth, TX: Botanical Research Institute of Texas (BRIT) Press. 779 p.
Macior, L.W. 1970. The pollination ecology of Pedicularis in Colorado. American Journal of Botany. 57(6): 716-728.
Maguire, A.D.; Overland, A. 1959. Laboratory germination of seeds of weedy and native plants. Washington State Agricultural Experiment Station Circular 349. Pullman, WA: Washington State University. 15 p.
Mansfield, D.H. 2000. Flora of Steens Mountain. Corvallis, OR: Oregon State University Press. 410 p.
McCulloch, C.Y. 1973. Part I: Seasonal diets of mule and white-tailed deer. In: Deer nutrition in Arizona chaparral and desert habitats. Special Report No. 3: Federal Aid in Wildlife Restoration Act Project W-78-R. Phoenix, AZ: Arizona Game and Fish Department, Research Division: 1-37.
Moerman, D. 2003. Native American ethnobotany: A database of foods, drugs, dyes, and fibers of Native American peoples, derived from plants. Dearborn, MI: University of Michigan. http://naeb.brit.org
Munz, P.A.; Keck, D.D. 1973. A California flora and supplement. Berkeley, CA: University of California Press. 1905 p.
Omernik, J.M. 1987. Ecoregions of the conterminous United States. Map (scale 1:7,500,000). Annals of the Association of American Geographers. 77(1): 118-125.
Parkinson, H.; DeBolt, A. 2005. Propagation protocol for production of container (plug) Chaenactis douglasii (Hook.) Hook. & Arn. plants. Native Plant Network. U.S. Department of Agriculture, Forest Service, National Center for Reforestation, Nurseries, and Genetic Resources. http://NativePlantNetwork.org [Accessed 2017 April 23].
Pavlik, B.M.; Ferguson, N.; Nelson, M. 1993. Assessing limitations on the growth of endangered plant populations, II. Seed production and seed bank dynamics of Erysimum capitatum ssp. angustatum and Oenothera deltoides ssp. howellii. Biological Conservation. 65(3): 267-278.
Pavlik, B.M.; Manning, E. 1993. Assessing limitations on the growth of endangered plant populations, I. Experimental demography of Erysimum capitatum ssp. angustatum and Oenothera deltoides ssp. howellii. Biological Conservation. 65(3): 257-265.
Pavlik, D.T.; Fleishman, E.; Wang, N.; Boswell, P.; Blair, R.B. 2018. Sugars in nectar sources and their use by butterflies (Hesperioidea and Papilionoidea) in the Sierra Nevada, California. Journal of the Lepidopterists’ Society. 72(2): 165-174.
Pearson, L.C. 1965. Primary production in grazed and ungrazed desert communities of eastern Idaho. Ecology. 46(3): 278-285.
Phillips, A.M. 1975. Flora of the Rampart Cave Area, Lower Grand Canyon, Arizona. Journal of the Arizona Academy of Science. 10(3): 148-159.
Plant Conservation Alliance [PCA]. 2015. National seed strategy for rehabilitation and restoration 2015-2020. Washington, DC: U.S. Department of the Interior, Bureau of Land Management. 52 p.
Plants for a Future [PFAF]. 2024. Available: https://pfaf.org/user/Plant.aspx
Preston, R.E. 1986. Pollen-ovule ratios in the Cruciferae. American Journal of Botany. 73(12): 1732-1740.
Price, R.A. 1986. Systematics of the Erysimum capitatum alliance (Brassicaceae). American Journal of Botany. 73(5): 781-781.
Richardson, B.; Kilkenny, F.; St. Clair, B.; Stevenson-Molnar, N. 2020. Climate Smart Restoration Tool. https://climaterestorationtool.org/csrt/
Robinson, G.S.; Ackery, P.R.; Kitching, I.; Beccaloni, G.W.; Hernández, L.M. 2023. HOSTS: A Database of the World’s Lepidopteran Hostplants [Data set]. Natural History Museum. https://doi.org/10.5519/havt50xw
Rossbach, G.B. 1958. New taxa and new combinations in the genus Erysimum in North America. Aliso. 4(1): 115-124.
Rundel, P.W.; Newman, M.; Rabenold, P. 2009. Plant communities and floristic diversity of the Emerald Lake Basin, Sequoia National Park, California. Madrono. 56(7): 184-198.
SEINet – Regional Networks of North American Herbaria Steering Committee [SEINet]. 2020. SEINet Regional Networks of North American Herbaria. https://Symbiota.org/docs/seinet
Sparrowhawk Native Plants. 2024. https://sparrowhawknativeplants.com
Spellenberg, R. 2012. Sonoran Desert wildflowers: A guide to common plants. Helena, MT: Falcon Guides. 245 p.
Stark, E.M. 2020. Pacific Northwest native plant profile: Western wallflower (Erysimum capitatum). https://realgardensgrownatives.com/ [Accessed 2024 July 17].
Stevens, J.T.; Miller, J.E.D.; Fornwalt, P.J. 2019. Fire severity and changing composition of forest understory plant communities. Journal of Vegetation Science. 30(6): 1099-1109.
Swank, G.R. 1932. The ethnobotany of the Acoma and Laguna Indians. Albuquerque, NM: University of New Mexico. Thesis. 86 p.
Taylor, R.J. 1992. Sagebrush country: A wildflower sanctuary. Missoula, MT: Mountain Press Publishing Company. 211 p.
Toliver, M.E. 2022. Observations on Euchloe olympia (W. H. Edwards) (Pieridae) in a central Illinois sand prairie. The Journal of the Lepidopterists’ Society. 74(2): 106-120.
Turkowski, F.J. 1975. Dietary adaptability of the desert cottontail. The Journal of Wildlife Management. 39(4): 748-756.
Turner, N.J.; Bouchard, R.; Kennedy, D.I. 1980. Ethnobotany of the Okanagan-Colville Indians of British Columbia and Washington. Victoria, BC: British Columbia Provincial Museum. 179 p.
USDA Forest Service, Bend Seed Extractory [USDA FS BSE]. 2021. Nursery Management Information System Version 4.1.11. Local Source Report 34-Source Received. Bend, OR: U.S. Department of Agriculture, Forest Service, Bend Seed Extractory.
USDA Forest Service, Western Wildland Environmental Threat Assessment Center [USFS WWETAC]. 2024. TRM Seed Zone Applications. Prineville, OR: U.S. Department of Agriculture, Forest Service, Western Wildland Environmental Threat Assessment Center. https://research.fs.usda.gov/pnw/products/dataandtools/seed-zone-webmap
USDA Natural Resources Conservation Service [USDA NRCS]. 2024. The PLANTS Database. Greensboro, NC: U.S. Department of Agriculture, Natural Resources Conservation Service, National Plant Data Team. https://plants.sc.egov.usda.gov/
USDI Bureau of Land Management, Seeds of Success [USDI BLM SOS]. 2021. Bureau of Land Management technical protocol for the collection, study, and conservation of seeds from native plant species for Seeds of Success. Washington, DC: U.S. Department of the Interior, Bureau of Land Management, Plant Conservation and Restoration Program. 44 p.
USDI Bureau of Land Management, Seeds of Success [USDI BLM SOS]. 2023. Bureau of Land Management technical protocol for the collection, study, and conservation of seeds from native plant species for Seeds of Success. Washington, DC: U.S. Department of the Interior, Bureau of Land Management, Plant Conservation and Restoration Program. 45 p.
USDI Environmental Protection Agency [USDI EPA]. 2018. Ecoregions. Washington, DC: U.S. Environmental Protection Agency. https://www.epa.gov/eco-research/ecoregions
USDI Geological Survey [USDI USGS]. 2020. Biodiversity Information Serving Our Nation (BISON). U.S. Geological Survey. https://www.gbif.us/
Utah Crop Improvement Association [UCIA]. 2015. How to be a seed connoisseur. Logan, UT: UCIA, Utah Department of Agriculture and Food, Utah State University and Utah State Seed Laboratory. 16 p.
Vestal, P.A. 1952. Ethnobotany of the Ramah Navaho. Reports of the Ramah Project: No. 4. Papers of the Peabody Museum of American Archeology and Ethnology. Cambridge, MA: Harvard University. 40(4): 1-94.
Vetter, M.A. 2000. Grasslands of the Aishihik-Sekulmun Lakes Area, Yukon Territory, Canada. Arctic. 53(2): 165-173.
Vogl, R.J.; Schorr, P. 1972. Fire and manzanita chaparral in the San Jacinto Mountains, California. Ecology. 53(6): 1179-1188.
Whittaker, R.H. 1960. Vegetation of the Siskiyou Mountains, Oregon and California. Ecological Monographs. 30(3): 279-338.
Wright, R.D.; Mooney, H.A. 1965. Substrate-oriented distribution of bristlecone pine in the White Mountains of California. The American Midland Naturalist. 73(2): 257-284.
Young, S.A.; Schrumpf, B.; Bouck, M.; Moore, M. 2020. How the Association of Official Seed Certifying Agencies (AOSCA) tracks wildland sourced seed and other plant propagating materials. Logan, UT: Utah State University, Utah Agricultural Experiment Station. 36 p.
How to Cite
Gucker, Corey L.; Shaw, Nancy L. 2024. Sanddune wallflower (Erysimum capitatum [Douglas ex Hook] Greene). In: Gucker, C.L.; Shaw, N.L., eds. Western forbs: Biology, ecology, and use in restoration. Reno, NV: Great Basin Fire Science Exchange. Online: https://westernforbs.org/species/sanddune-wallflower-erysimum-capitatum/