Authorship
Gucker, Corey; Shaw, Nancy
Publication Date
September 2023
Nomenclature
Largeflower hawksbeard belongs to the Asteraceae family and the Psilochaenia section of the Crepis genus (Sears and Whitton 2016).
Family
Asteraceae – Aster family
Genus
Crepis
Species
occidentalis
NRCS Plant Code
CROC (USDA NRCS 2023).
Subtaxa
The Flora of North America (Bogler 2006) recognizes four subspecies: C. o. subsp. conjuncta, C. o. subsp. costata, C. o. subsp. occidentalis, and C. o. subsp. pumila.
Synonyms
Psilochenia occidentalis (Nuttall) Nuttall (Bogler 2006).
Common Names
Largeflower hawksbeard and western hawksbeard (Andersen and Holmgren 1996).
Chromosome Number
Largeflower hawksbeard is a polymorphic species with chromosome numbers of 2n = 22, 33, 44, 55, 66, 77, and 88 (Hickman 1993; Cronquist 1994; Bogler 2006). Chromosome numbers vary by subspecies. Chromosome numbers for conjuncta and costata are 2n = 44, 77 or 88, occidentalis 2n = 22, 33, 44, and pumila 2n = 33, 55, 77 (Babcock and Stebbins 1938).
Hybridization
There is considerable hybridization in the Crepis genus and largeflower hawksbeard subspecies intergrade extensively (Babcock and Stebbins 1938; Hickman 1993). Subspecies conjuncta reflects past hybridization between Modoc hawksbeard (C. modocensis) and Baker’s hawksbeard (C. bakeri) (Cronquist 1994).
Distribution
Largeflower hawksbeard has a widespread western distribution from British Columbia to Saskatchewan in the north to South Dakota in the east and in New Mexico, Arizona, and California in the south (Hermann 1966; Bogler 2006). Populations are most common in the Columbia Plateau and Great Basin and frequent in northern California and southern Oregon (Babcock and Stebbins 1938; Sears and Whitton 2016).
Subspecies occidentalis occurs throughout the species range (Bogler 2006). Subspecies conjuncta occurs in Washington, Montana, Wyoming, Colorado, California, and Oregon. Subspecies costata occurs in British Columbia, Saskatchewan, Montana, Wyoming, Colorado, Utah, Nevada, California, Oregon, Washington, and Idaho. Subspecies pumila occurs in British Columbia, Washington, Oregon, Idaho, Montana, Utah, Nevada, and California (Bogler 2006).
Babcock and Stebbins (1938) suggest that largeflower hawksbeard is one of the most widespread and perhaps the most polymorphic of the American hawksbeards (Crepis spp.). In a thorough study of the species, they mapped the distribution of the polyploid and apomictic forms (Fig. 1). The diploid sexual form has a very restricted distribution, occurring at elevations of 3,900 to 5,200 ft (1,200-1,600 m) on the arid east side of the Sierra Nevada Range in northeastern California and adjacent Nevada. Polyploid forms, especially those with chromosome numbers of 2n = 33 or 44 occurred nearly throughout the species range. Polyploids with higher chromosome numbers were less widespread and tended to be more abundant near the centers of the subspecies’ distributions (Babcock and Stebbins 1938).
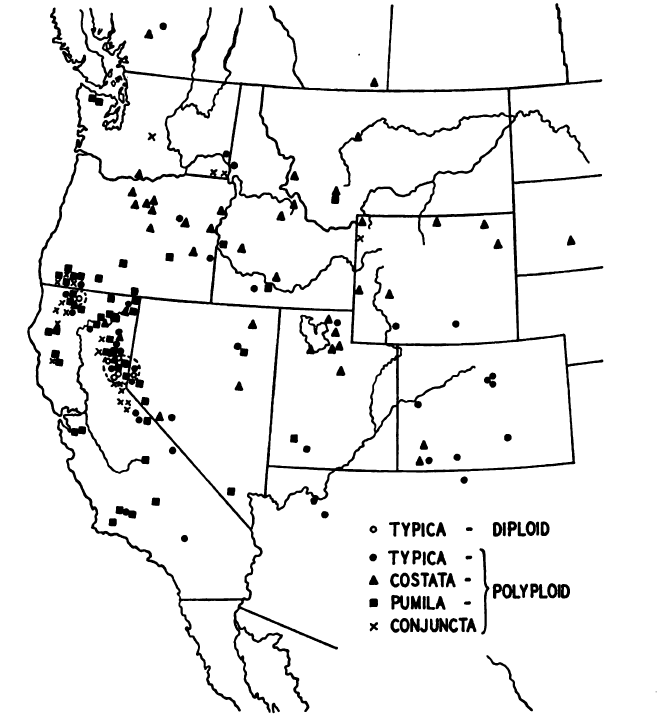
Figure 1. Distribution of largeflower hawksbeard and its sexual diploid (subsp. occidentalis [typica]) and apomictic polyploid forms (subsps. occidentalis [typica in figure], conjuncta, costata, and pumila) (from Babcock and Stebbins 1938).
Habitat And Plant Associations
Largeflower hawksbeard grows on dry, rocky open slopes, and some of the most arid sites occupied by American hawksbeards in the West included largeflower hawksbeard populations (Babcock and Stebbins 1938). Largeflower hawksbeard commonly occurs in bunchgrass (Fig. 2), sagebrush (Artemisia spp.) (Fig. 3), and pinyon-juniper (Pinus–Juniperus spp.) communities (Babcock and Stebbins 1938; Hickman 1993; Cronquist 1994; Blackwell 2006; Hitchcock and Cronquist 2018). It also grows in mountain brush, quaking aspen (Populus tremuloides), shadscale saltbush (Atriplex confertifolia), rabbitbrush (Chrysothamnus spp.), ponderosa pine (Pinus ponderosa), and lodgepole pine (P. contorta) vegetation (Munz and Keck 1973; Welsh et al. 2016). Subspecies conjuncta grows in forested areas with higher precipitation levels than the vegetation types providing habitat to subspecies costata, which is common in juniper-oak (Quercus spp.) woodlands, or subspecies occidentalis, which is common in sagebrush scrub, or subspecies pumila, which is common in open pine (Pinus spp.) woodlands (Babcock and Stebbins 1938; Bogler 2006).

Figure 2. Largeflower hawksbeard growing in a grassland in Utah. Photo: USDA, Forest Service, Provo Shrub Sciences Laboratory.

Figure 3. Dense population of largeflower hawksbeard growing in sagebrush habitat in Idaho. Photo: USDI Bureau of Land Management (BLM) ID931, Seeds of Success (SOS).
Cover of largeflower hawksbeard reached up to 5% in the bluebunch wheatgrass-Sandberg bluegrass-onespike danthonia (Pseudoroegneria spicata–Poa secunda–Danthonia unispicata) vegetation type in a vegetation survey of the Blue and Ochoco Mountains in eastern Oregon. This vegetation type occurred primarily on southern aspects between elevations of 3,500 to 5,150 ft (1,070-1,570 m) (Johnson and Swanson 2005). In a survey of soil, plant, and climate relationships throughout sagebrush communities in the Intermountain West, largeflower hawksbeard was most common in Wyoming big sagebrush (Artemisia tridentata subsp. wyomingensis)/bluebunch wheatgrass communities. The study area had a continental climate with arid hot summers, cold winters, low humidity levels, and wide yearly and seasonal fluctuations in precipitation and temperature (Passey et al. 1982). Largeflower hawksbeard occurred in grazed and protected big sagebrush/needle and thread (Artemisia tridentata/Hesperostipa comata) plant communities at the northeastern end of the Snake River Plain near Rexburg, Idaho. This area experiences cold winters, hot summers, low average annual precipitation (10.6 in [270 mm]), high winds, and extreme temperature fluctuations (-44 to 104 °F [-42 to 40 °C]) (Pearson 1965).
In a floristic inventory of Montana’s Phillips and Valley Counties, largeflower hawksbeard occurred at elevations of 2,030 to 3,760 ft (620-1,145 m) in mixed-grass prairie, sagebrush steppe with Wyoming big sagebrush and silver sagebrush (A. cana subsp. cana), and ponderosa pine-Rocky Mountain juniper (Juniperus scopulorum) on steep drainages of the Missouri River Breaks (Charboneau et al. 2013). In a vegetation survey of the Wind River Range in northwestern Wyoming, largeflower hawksbeard (subsp. costata) occurred at elevations of 5,748 to 7,398 ft (1,752-2,255 m) in mountain big sagebrush (A. t. subsp. vaseyana) grasslands, juniper woodlands, and disturbed sites. A narrower elevation range of 5,659 to 6,283 ft (1,725-1,915 m) was reported for subsp. occidentalis in big sagebrush grasslands (Fertig et al. 2013).
Largeflower hawksbeard occurred in very low abundance in Utah juniper (J. osteosperma)/bluebunch wheatgrass, black sagebrush (A. nova)/bluebunch wheatgrass, Wyoming big sagebrush/bluebunch wheatgrass, and shadscale saltbush/bluebunch wheatgrass in a survey of vegetation in northwestern Colorado. This vegetation survey focused on remnants unaltered by grazing, logging, or other human-related uses (Baker and Kennedy 1985). In the Piceance Basin, Colorado, largeflower hawksbeard constancy was 33% in Douglas-fir/Utah serviceberry-Gambel oak (Pseudotsuga menziesii/Amelanchier utahensis–Q. gambelii) vegetation on north slopes at elevations of 6,230 to 8,530 ft (1,900-2,600 m). Constancy was 67% in Wyoming big sagebrush-mountain snowberry/basin wildrye (Symphoricarpos oreophilus/Leymus cinereus) at elevations of 7,220 to 8,530 ft (2,200-2,600 m). In the Piceance Basin, annual precipitation averages 13 in (338 mm) and temperatures average 18 °F (-8 °C) in January and 67 °F (19 °C) in July (Baker 1983).
In Utah, largeflower hawksbeard occurred in Gambel oak woodlands (Kunzler et al. 1981) and curl-leaf mountain mahogany (Cercocarpus ledifolius) communities (Davis 1976). In Nevada, it occurred at elevations of 5,800 to 7,000 ft (1,800-2,100 m) in sagebrush and pinyon-juniper vegetation (Beatley 1976). In a survey of upper elevations of the Kaibab Plateau in Arizona’s Coconino County, largeflower hawksbeard occurred in pine forests (Rink et al. 2020). In a vegetation survey of the mountains in north-central New Mexico, largeflower hawksbeard occurred in montane shrublands at 8,200 to 8,500 feet (2,500-2,600 m) in the Tusas Mountains. These shrublands occurred as patches in ponderosa pine forests and twoneedle pinyon-oneseed juniper (Pinus edulis–J. monosperma) woodlands. The shrublands were dominated by chokecherry (Prunus virginiana), Gambel oak, or alderleaf mountain mahogany (C. montanus) and were often indicative of past disturbance (Reif et al. 2009).
Elevation
Largeflower hawksbeard occurs at elevations from 2,000 to 9,020 ft (600-2,750 m) throughout its range (Munz and Keck 1973; Cronquist 1994; Blackwell 2006; Bogler 2006). The elevation range for subspecies conjuncta is 4,600 to 6,890 ft (1,400-2,100 m), for subspecies costata is 3,940 to 8,200 ft (1,200-2,500 m), for subpsecies occidentalis 3,280 to 7,220 ft (1,000-2,200 m), and for subspecies pumila 2,630 to 5,910 ft (800-1,800 m) (Bogler 2006). The elevation range in Utah is 5,000 to 7,220 ft (1,525-2,200 m) for subspecies costata, 4,200 to 8,420 ft (1,280-2,565 m) for subspecies occidentalis, and 5,580 to 6,890 ft (1,700-2,100 m) for subspecies pumila (Welsh et al. 2016).
Soils
Largeflower hawksbeard often occupies well-drained sites with deep, rich soils (Babcock and Stebbins 1938). Subspecies conjuncta and occidentalis commonly occur on gravelly or rocky soils (Fig. 4). Subspecies costata is often associated with black shale or sandstone, and subspecies pumila tolerates serpentine soils (Bogler 2006).

Figure 4. Small largeflower hawksbeard plants growing on a very rocky site in Oregon. Photo: BLM OR010, SOS.
Largeflower hawksbeard grows on a variety of soil textures. In an evaluation of 107 late seral and relatively undisturbed Wyoming big sagebrush habitats across southeastern Oregon and northern Nevada, largeflower hawksbeard was positively associated with soils having more clay and less sand in the top 6 in (15 cm) (Davies et al. 2007). In the Blue and Ochoco Mountains of eastern Oregon, it occurred in grasslands on basalt and volcanic tufts. The top 5 in (13 cm) of these soils were gravelly loams over very to extremely gravelly clay loams with the depth to bedrock ranging from 6 to 20 in (15-51 cm) (Johnson and Swanson 2005). Throughout Intermountain West sagebrush communities, largeflower hawksbeard was most common in Wyoming big sagebrush/bluebunch wheatgrass communities that occupied frigid to mesic soils ranging from fine, fine silty, coarse silty, clayey, to fine loamy (Passey et al. 1982). In north-central New Mexico, it grew in montane shrublands on rocky sites with poorly developed xeric soils (Reif et al. 2009).
Although deep soils are preferred, largeflower hawksbeard also occurs on shallow soils. On kipuka sites, areas where natural plant communities are isolated by lava flows, near Blackfoot, Idaho, largeflower hawksbeard occurred on lava and small basalt outcrops with roots concentrated in the top 12 in (30 cm) of soil (Passey and Hugie 1963). At other eastern Idaho sites, where largeflower hawksbeard grows with big sagebrush, soils were sandy, shallow (about 24 in [60 cm]), and underlain by lava rock. Soil pH was 7.8, low in humus and nitrogen, moderately high in phosphorus, and very high in potassium, sulphur, calcium, and magnesium (Pearson 1965). In southeastern Oregon and northeastern California where largeflower hawksbeard occurs in mountain big sagebrush habitats, soils were well to moderately well drained and 14 to 32 in (36-80 cm) deep (Miller et al. 2000).
Description
Largeflower hawksbeard is a highly variable, stout perennial from a caudex and a deep, thick taproot (Babcock and Stebbins 1938; Hickman 1993; Bogler 2006). The caudex is often covered by persistent withered brown leaf bases (Welsh et al. 2016). Taproots are very long, slender, and woody (Babcock and Stebbins 1938). Rooting depth averaged 8 in (20 cm) for largeflower hawksbeard plants growing in late-seral Idaho fescue (Festuca idahoensis)-bluebunch wheatgrass sites in southwestern Montana (Pokorney et al. 2004).
Plants produce one to three erect, candelabra-like main stems 3 to 16 in (8-40 cm) tall that branch from the middle or less often from near the base (Fig. 5) (Coville 1896; Babcock and Stebbins 1938; Munz and Keck 1973; Hickman 1993; Cronquist 1994; Blackwell 2006; Bogler 2006). Stem bases are typically yellow. Herbage contains a milky sap and is densely and closely gray-tomentose but becomes more glabrate with age (Babcock and Stebbins 1938; Munz and Keck 1973; Cronquist 1994; Lesica 2012; Welsh et al. 2016). The tomentose nature may also vary by habitat. Coville (1896) reported It is denser and more common tomentose characteristics for plants in dry exposed habitats and becomes less glandular-hirsute characteristics for plants in milder Pacific Coast climates.

Figure 5. Largeflower hawksbeard plant in flower. Notice that flower buds, open flowers, and faded flowers all occur on this single plant. Photo: BLM CA350, SOS.

Figure 6. Basal leaves of a largeflower hawksbeard plant growing in Idaho. Photo: BLM ID230, SOS.
Each main plant branch produces 2 to 30 broad flower heads (1.25 in [3 cm] wide) in a loose corymbiform inflorescence (Munz and Keck 1973; Cronquist 1994; Andersen and Holmgren 1996; Bogler 2006; Welsh et al. 2016). Individual flower heads include 10 to 40 perfect, yellow, ray florets (Hermann 1966; Cronquist 1994; Bogler 2006). Florets are 0.4 to 0.6 in (10-15 mm) long and have square tips (Blackwell 2006; Lesica 2012). Involucres are narrowly bell-shaped and 0.4 to 0.8 in (1-2 cm) high and 0.2 to 0.6 in (6-15 mm) wide with six to eight outer phyllaries, which are about half as long as the 8 to 13 inner phyllaries (Coville 1896; Babcock and Stebbins 1938; Munz and Keck 1973; Hickman 1993; Welsh et al. 2016; Hitchcock and Cronquist 2018; Luna et al. 2018). Phyllaries range from glandular to tomentose, sometimes with blackish to greenish gland-tipped bristles (Blackwell 2006; Hitchcock and Cronquist 2018).
Largeflower hawksbeard produces achenes, commonly and hereafter referred to as seeds. They are yellow brown to dark brown, subcylindric, and 6 to 10 mm long (Cronquist 1994; Bogler 2006). Seed ends are tapered, and the body is moderately to strongly 10 to 18 ribbed (Babcock and Stebbins 1938; Munz and Keck 1973). Each seed has a copious, soft, dusky white to yellowish white pappus that is 10 to 12 mm long and exceeds the involucre at maturity (Babcock and Stebbins 1938; Hickman 1993; Bogler 2006).
Distinguishing Subspecies.
Subspecies conjuncta is 2 to 8 in (5-20 cm) tall with stems that branch near their base (Babcock and Stebbins 1938; Munz and Keck 1973; Cronquist 1994; Bogler 2006). Herbage is tomentose. Leaves are 4 to 7 in (10-18 cm) long, 1.2 to 1.6 in (3-4 cm) wide, and range from pinnatifid or deeply lobed to few toothed or entire. Plants produce two to nine flower heads, each often comprised of less than 15 florets. Involucres have glandular pubescence, 8 to 12 inner bracts with outer bracts (5-9 mm) being half to two-thirds as long as the inner bracts (Babcock and Stebbins 1938; Munz and Keck 1973; Cronquist 1994; Bogler 2006). Seeds are dark brown (Bogler 2006).
Subspecies costata is 3 to 16 in (8-40 cm) tall with tomentose herbage. The involucres, peduncles, and upper stem leaves often have conspicuous, relatively large, dark to black, gland-tipped trichomes (Babcock and Stebbins 1938; Munz and Keck 1973; Cronquist 1994; Bogler 2006). Leaves are 2 to 8 in (5-20 cm) long, 0.8 to 1.4 in (2-3.5 cm) wide, and pinnately dentate lobed. Plants often have 15 to 30 flower heads, typically comprised of 10 to 14 florets. Involucres have 7 to 8 inner bracts (Babcock and Stebbins 1938; Munz and Keck 1973; Cronquist 1994; Bogler 2006). Seeds are golden brown (Bogler 2006).
Polyploids of subspecies occidentalis are not always morphologically distinguishable from diploids of subspecies occidentalis, but they can almost always be distinguished by stomata size and pollen grain morphology (Babcock and Stebbins 1938). Subspecies occidentalis is 4 to 16 in (10-40 cm) tall with slightly to strongly stipitate glandular herbage (Babcock and Stebbins 1938; Munz and Keck 1973; Cronquist 1994; Bogler 2006). Leaves are 4 to 8 in (10-20 cm) long, 0.8 to 1.6 in (2-4 cm) wide, and sharply dentate to pinnately lobed. Plants often have 10 to 30 flower heads, which are comprised of 18 to 30 florets. Involucres have 10 to 13 inner bracts Babcock and Stebbins 1938; Munz and Keck 1973; Cronquist 1994; Bogler 2006). Seeds are golden brown (Bogler 2006).
Subspecies pumila is 4 to 16 in (10-40 cm) tall with well-developed stems having definite primary axes and distal branching (Babcock and Stebbins 1938; Munz and Keck 1973; Cronquist 1994; Bogler 2006). Herbage is tomentulose but the pubescence is non glandular. Leaves are 4 to 8 in (10-20 cm) long, 0.8 to 2 in (2-5 cm) wide, and coarsely dentate to pinnately lobed. Plants often have 5 to 15 flower heads of 12 to 20 florets. Involucres typically have 8 inner bracts with the outer bracts being a third to half as long as the inner bracts (Babcock and Stebbins 1938; Munz and Keck 1973; Cronquist 1994; Bogler 2006). Seeds are brown (Bogler 2006).
Reproduction
Largeflower hawksbeard reproduces from seed (Fig. 7), which is produced sexually in diploid plant forms and asexually through mostly or wholly apomixis in polyploid forms (Babcock and Stebbins 1938; Stebbins and Jenkins 1939). In a study evaluating the patterns of co-occurrence of sexual and apomictic hawksbeards, researchers found no obvious habitat distinctions between places where sexual and apomictic forms grow. Habitats were not distinct although distributions were for the sexual and apomictic forms (see Distribution). Most apomictic forms produced some viable pollen, which provides an opportunity for reproductive interference by reducing the reproductive success of nearby sexual relatives and potential local exclusion of sexual forms (Whitton et al. 2017).
Pollination
Flowers are self-incompatible (Stebbins and Jenkins 1939), so pollination is necessary for seed production. Specific pollinator relationships were not described in the reviewed literature, but the plants do attract bees (Tilley et al. 2013). Several bee genera were collected from largeflower hawksbeard flowers from just a handful of sagebrush sites in eastern Oregon (S. Copeland, USDA ARS, personal communication, November 2023).
Flowers are typically produced from May to July (LBJWC 2022) although they can occur as early as April and as late as August (Munz and Keck 1973; Beatley 1976). Subspecies occidentalis produces flowers a little earlier than subspecies conjuncta, costata, and pumila (Bogler 2006).
Ecology
Hawksbeard species are somewhat long-lived since they take 3 to 5 years to reach reproductive maturity (Jensen et al. 2011).
Largeflower hawksbeard occurs on disturbed (Fertig et al. 2013) and grazed sites (Pearson 1965). It recovers rapidly, and often to beyond predisturbance levels, after fire (Bates et al. 2007) and after tree removal and fire (Bates and Svejcar 2009).
Seed And Seedling Ecology
Insect larvae predation of seeds and seedheads is common for largeflower hawksbeard (Babcock and Stebbins 1938).

Figure 7. Largeflower hawksbeard plant beginning to produce seed. Photo: BLM CA170, SOS.
Studies suggest that largeflower hawksbeard seedlings grow slowly even under ideal conditions (James and Drenovsky 2007). In the greenhouse, the relative growth rate (RGR) of seedlings averaged 0.08 g/g/day at 14 days, 0.06 g/g/day at 28 days, and 0.04 g/g/day at 42 days after the appearance of cotyledons. Largeflower hawksbeard’s RGR and was lowest among the native forbs evaluated (Munro’s globemallow [Sphaeralcea munroana]*, common yarrow [Achillea millefolium]*, common woolly sunflower [Eriophyllum lanatum]*, Rocky Mountain penstemon [Penstemon strictus], and Lewis flax [Linum lewisii], listed in order of decreasing RGR and *indicating significantly greater than RGR for largeflower hawksbeard). Largeflower hawksbeard RGR was significantly lower than that of the invasive species evaluated, which had RGRs ranging from 0.11 to 0.15 g/g/day at 42 days (spotted knapweed [Centaurea stoebe], rush skeletonweed [Chondrilla juncea], Dalmatian toadflax [Linaria dalmatica], whitetop [Cardaria draba], Fuller’s teasel [Dipsacus fullonum], and Scotch cottonthistle [Onopordum acanthium]). Largeflower hawksbeard also had the lowest relative root elongation rate of all native and nonnative species evaluated, but the differences were not significant. This study was conducted in a greenhouse at the Eastern Oregon Agricultural Research Center, USDA ARS in Burns, Oregon. Daily average greenhouse temperatures were 46 °F (8 °C) for the low and 84 °F (29 °C) for the high. Seeds were planted in 1:1 mix of sandy loam field soil and coarse sand (James and Drenovsky 2007).
Disturbance Ecology
Largeflower hawksbeard tolerates disturbance and abundance is often greater on grazed or burned than protected or unburned sites (Pearson 1965; Ott et al. 2001, 2003; Bates and Svejcar 2009).
Frequency of largeflower hawksbeard was greater on grazed than on protected big sagebrush sites at the northeastern end of the Snake River Plain. Frequency averaged 5% in plots that had been protected from grazing by dairy cattle for 11 years and 18% in plots grazed for about 70 years. Exclosures were installed and protected at both sites during the 2-year study (Pearson 1965).
Largeflower hawksbeard was not affected or had increased abundance after fire in big sagebrush grassland vegetation in Oregon (Sapsis 1990; Bates et al. 2011). Plants were unharmed by a low-severity, growing-season prescribed fire (May 24, 1988) in basin big sagebrush/Idaho fescue-bluebunch wheatgrass communities at the John Day Fossil Bed National Monument, eastern Oregon. The frequency of largeflower hawksbeard was 10% before and 9% in post-fire year 1 (Sapsis 1990). The cover of largeflower hawksbeard increased by up to 4 times prefire levels following an August wildfire in Wyoming big sagebrush vegetation in the Sheepshead Mountains of southeastern Oregon. The wildfire removed most sagebrush shrubs from the mid- to late-seral stands. Recovery of largeflower hawksbeard was considered good in postfire monitoring that occurred for 4 post-fire years (Bates et al. 2007). Following a fall prescribed fire in late-seral Wyoming big sagebrush on the Northern Great Basin Experimental Range about 62 mi (100 km) southeast of Burns, Oregon, largeflower hawksbeard was listed among those forbs unaffected or having greater cover on burned than unburned plots. The fall prescribed fire killed 92% of Wyoming big sagebrush shrubs (Bates et al. 2011).
The cover of largeflower hawksbeard was greater in cut and burned than in cut and unburned plots for up to 3 years following treatments in western juniper (Juniperus occidentalis)-dominated sites on Steens Mountain, southeastern Oregon. Prior to the treatments, plots averaged 31% cover of trees that were 90 years old. Cutting occurred in 1998, burning occurred in 1998 or 1999. By 2006, largeflower hawksbeard abundance differences were no longer apparent on treated plots (Bates and Svejcar 2009).
Largeflower hawksbeard tolerated fire and post-fire revegetation efforts in big sagebrush, Utah juniper, and pinyon-juniper sites in Utah (Ott et al. 2001, 2003). Wildfires occurred in July or August 1996. Frequency of largeflower hawksbeard was greater on burned than unburned plots, and differences in frequency remained stable over 3 years of postfire monitoring. Researchers considered largeflower hawksbeard an early seral species because of the abundance differences between burned and unburned plots (Ott et al. 2001). Burned areas were aerially seeded with a mix of nonnative grasses, forbs, and fourwing saltbush (Atriplex canescens). Largeflower hawksbeard was not seeded but occurred on treated sites. Frequency of largeflower hawksbeard was lower on plots where seeding was followed by chaining (Ely chain) than on seeded plots that were not chained. Frequency differences on the revegetated plots remained for 3 years of posttreatment monitoring (Ott et al. 2003).
Wildlife And Livestock Use
Largeflower hawksbeard provides good forage for large ungulates, livestock, and game birds, and attracts bees (Hermann 1966; Mackie 1970; Murray et al. 1978; Miller et al. 1981; Tilley et al. 2013).
Hawksbeards in general and largeflower hawksbeard specifically are eaten by elk (Cervus canadensis), deer (Odocoileus spp.), and pronghorn (Antilocapra americana). Hawksbeards are considered high-value elk forage (Geary et al. 2017). Habitat use and diets of deer and elk were evaluated from mid-March to late October in the foothills of the Wallowa Mountains in northeastern Oregon. Largeflower hawksbeard was 1% of big game diets but had a very high preference index of 80. The preference index was determined by dividing the amount of largeflower hawksbeard in diets by its amount in the study area (Miller et al. 1981). In the Missouri River Breaks in northeastern Fergus County, Montana, heavy use of largeflower hawksbeard by mule deer in late May was noted at some sites (Mackie 1970). In central Montana, largeflower hawksbeard was 3% of June pronghorn diets in summer feeding observations. Feeding was observed from June through August for 2 years in various vegetation types. Largeflower hawksbeard occurred in big sagebrush/grassland vegetation which made up 13 or 14 of the 62 feeding sites visited each year (Wentland 1968).
Largeflower hawksbeard is good forage for livestock, especially sheep (Hermann 1966). At Saylor Creek Experimental Range, cattle readily fed on initial growth of largeflower hawksbeard in late spring and regrowth in late summer (Murray et al. 1978).
Largeflower hawksbeard plant material was recovered from a packrat (Neotoma spp.) midden in the Virginia Mountains of northwestern Nevada. It was recovered from the midden at present day, suggesting that largeflower hawksbeard is used by packrats in nest construction or as food (Nowak et al. 1994).
Asteraceae containing milky sap and hawksbeard species are eaten by greater sage-grouse (Centrocercus urophasianus) (Pennington et al. 2016; Luna et al. 2018) and are considered a highly valuable food (see Table 1, Gregg et al. 2008). In a review on the importance of forbs in diets, it was noted that greater sage-grouse females in Oregon and Nevada commonly consumed hawksbeards, which were reported in the habitats or identified as a food source in 7 of the 68 sources reviewed (Pennington et al. 2016). Researchers collected and examined the crops of 86 preincubating females in southeastern Oregon and northwestern Nevada and found that hawksbeards were among the most frequent forbs in sage-grouse crops (>15%) from mid-March to mid-April (Gregg et al. 2008). Use of hawksbeards increased with increasing annual availability of forbs, which coincided with this period. Hawksbeards made up a high of 23% of the dry mass of adult greater sage-grouse diets (Gregg et al. 2008).
Nutritional Value
Nutrient content of hawksbeards consumed by preincubating female greater sage-grouse is provided in Table 1. Plant samples were collected from foraging areas in southeastern Oregon and northwestern Nevada in March and April (Gregg et al. 2008). Oil content of seeds (not consumed by greater sage-grouse) averages 22% (SER, INSR, RBGK 2023).
Table 1. Nutrient content of hawksbeard plants collected from greater sage-grouse foraging areas in southeastern Oregon and northwestern Nevada for two years in March and April (Gregg et al. 2008).
| Year | Crude protein | Calcium | Phosphorus |
| ——————–%—————— | |||
| 2002 | 31 | 0.98 | 0.61 |
| 2003 | 26 | 0.77 | 0.53 |
Ethnobotany
The Northern Paiute people of the Surprise Valley area of California ate largeflower hawksbeard leaves raw as greens. Stems and roots were not used (Kelly 1932; Gamble 2008).
Horticulture
Introduction of hawksbeard species into horticulture is difficult because seedlings grow slowly, are sensitive to excess moisture and poor drainage, and produce long taproots. As of 1950, hawksbeard plants had not been grown to the flowering stage in cultivation (Stebbins 1950 cited in Sears and Whitton 2016). Based on more recent personal experience, Sears and Whitton (2016) report that propagation difficulties begin at the seed germination and establishment phases.
Revegetation Use
Largeflower hawksbeard has a moderate lifespan, tolerates disturbance, attracts bees, and is recommended for revegetation where annual precipitation averages 12 to 18 in (305-457 mm) (Eldredge et al. 2013; Fertig et al. 2013; Tilley et al. 2013). It is recommended for all Major Land Resource Areas in Nevada (Eldredge et al. 2013).
Developing A Seed Supply
For restoration to be successful, the right seed needs to be planted in the right place at the right time. Coordinated planning and cooperation is required among all partners to select appropriate species and seed sources; these are determined by current or projected revegetation requirements and goals (PCA 2015).
Developing a seed supply begins with seed collection from native stands. Collection sites are determined by current or projected revegetation requirements and goals as well as through consultation with land managers and stakeholders. Collected seed is distributed for restoration or first increased in seed fields to provide a larger supply of seed (PCA 2015). Production of nursery stock requires less seed than large-scale seeding operations. Regardless of the size and complexity of any revegetation effort, seed certification is essential for tracking seed origin from collection through use (UCIA 2015).
Seed Sourcing
Because empirical seed zones are not currently available for largeflower hawksbeard, generalized provisional seed zones developed by Bower et al. (2014) may be used to select and deploy seed sources. These provisional seed zones identify areas of climatic similarity with comparable winter minimum temperature and aridity (annual heat to moisture index). In Figure 8, Omernik Level III Ecoregions (Omernik 1987) overlay the provisional seed zones to identify climatically similar but ecologically different areas. For site-specific disturbance regimes and restoration objectives, seed collection locations within a seed zone and ecoregion may be further limited by elevation, soil type, or other factors.
The Western Wildland Environmental Threat Assessment Center’s (USDA FS WWETAC 2017) Threat and Resource Mapping (TRM) Seed Zone application provides links to interactive mapping features useful for seed collection and deployment planning. The Climate Smart Restoration Tool (Richardson et al. 2020) can also guide revegetation planning, seed collection, and seed deployment, particularly when addressing climate change considerations.
Occurrence Map
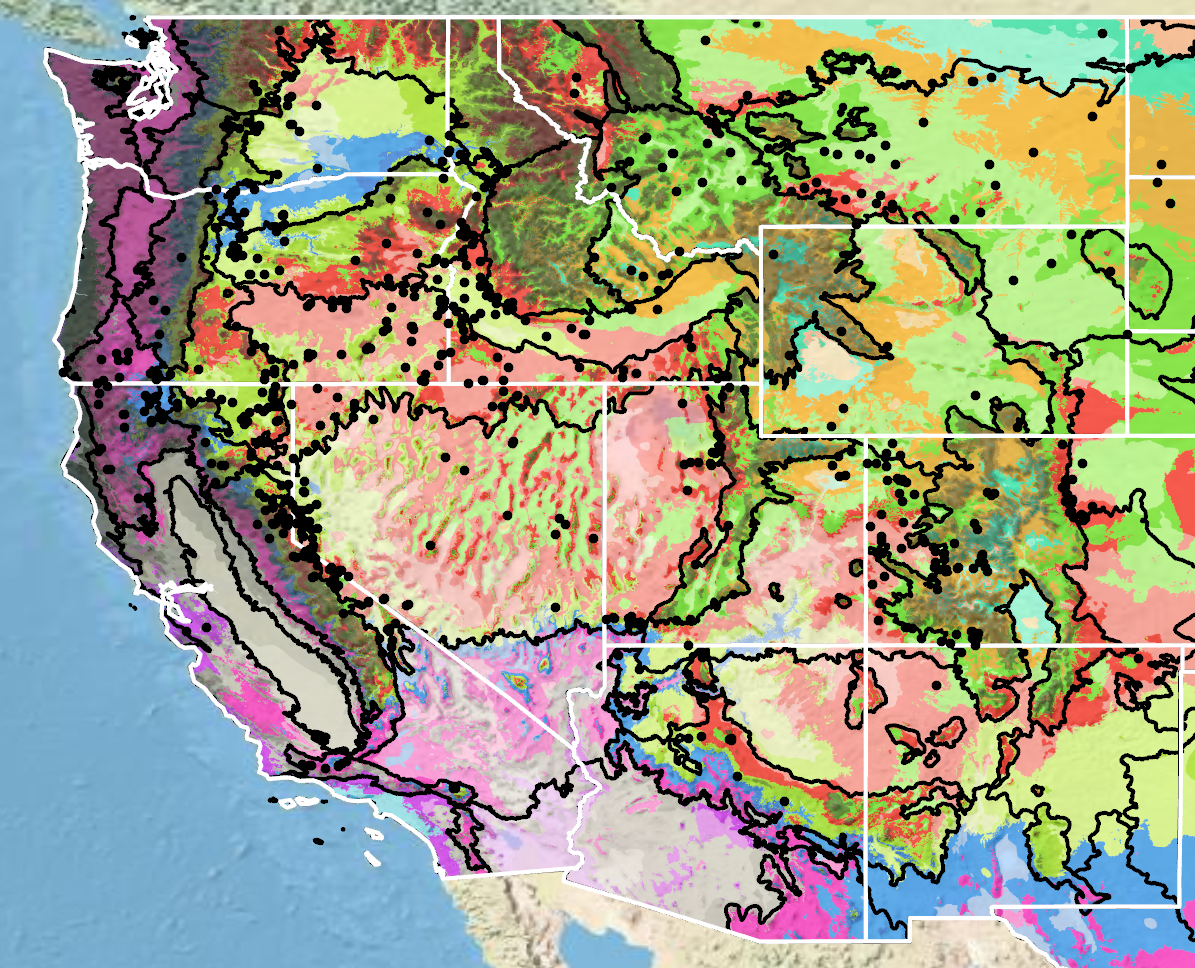
Figure 8. Distribution of largeflower hawksbeard (black circles) based on geo-referenced herbarium specimens and observational data from 1865-2023 (CPNWH 2017; SEINet 2020; USDI USGS 2020). Generalized provisional seed zones (colored regions) (Bower et al. 2014) are overlain by Omernik Level III Ecoregions (black outlines) (Omernik 1987; USDI EPA 2018). Interactive maps, legends, and a mobile app are available (USDA FS WWETAC 2017; research.fs.usda.gov/pnw/products/dataandtools/seed-zone-webmap). Map prepared by S. Barga and B. Bautch-Breitung, USDA Forest Service, RMRS.
Releases
As of 2023, there were no largeflower hawksbeard germplasm releases.
Wildland Seed Collection
Seed is generally ripe once the dusky white pappus is visible in the seedhead (Fig. 9), which often occurs in June or July (USDI BLM SOS 2020). Viable wildland seed is typically dark and firm (Fig. 10), but insect damage is common (Babcock and Stebbins 1938). Viable seeds are visibly wider than nonviable seeds (Wirth 2000).

Figure 9. Largeflower hawksbeard seedheads with ripening seed. Photo: BLM NV030, SOS.
Wildland Seed Certification
Verification of species and tracking of geographic source is necessary whether wildland seed is collected for immediate project use or as stock seed for cultivated increase. This official Source Identification process can be accomplished by following procedures established by the Association of Official Seed Certifying Agencies (AOSCA) Pre-Variety Germplasm Program (UCIA 2015; Young et al. 2020). Wildland seed collectors should become acquainted with state certification agency procedures, regulations, and deadlines in the states where they collect.
If wildland-collected seed is to be sold for direct use in ecological restoration projects, collectors must apply for Source-Identified certification prior to making collections. Pre-collection applications, site inspections, and species and seed amount verification are handled by the AOSCA member state agency where seed collections will be made (see listings at AOSCA.org).
If wildland seed collected by a grower or private collector is to be used as stock seed for planting cultivated seed fields or for nursery propagation (See Agricultural Seed Field Certification section), detailed information regarding the collection site and collecting procedures must be provided when applying for certification. Photos and herbarium specimens may be required. Germplasm accessions acquired within established protocols of recognized public agencies, however, are normally eligible to enter the certification process as stock seed without routine certification agency site inspections. For contract grow-outs, however, this collection site information must be provided to the grower to enable certification.
Collection Timing
The USDI Bureau of Land Management’s Seeds of Success field crews provided data from 31 largeflower hawksbeard collections. Harvests were made in 9 years from 2002 to 2016 in California (4 collections), Oregon (12), Idaho (7), Nevada (5), Utah (2), and Wyoming (1). The earliest harvest date was June 4, 2013, in Washington County, Idaho, at an elevation of 2,208 ft (673 m). The latest harvest date was July 21, 2011, in Washoe County, Nevada, at an elevation of 6,108 ft (1,862 m). At four sites more than one collection was made. Subsequent collections were made within 2 days and up to 2 weeks of previous collections. In the single year with the most collections made, eight in 2010, the earliest collection was June 24 at 4,804 ft (1,464 m) elevation in Carson City County, Nevada. The latest collection date in 2010 was July 20 in Gooding County, Idaho, at 3,150 ft (960 m), suggesting that factors other than elevation are important to the timing of seed maturation (USDI BLM SOS 2020).
Collection Methods
Wildland seed can be collected by hand-stripping, using racquets to swat the seed heads over a collection hopper (St. John and Tilley 2012) or by clipping the seed heads (Fig. 8).
Several collection guidelines and methods should be followed to maximize the genetic diversity of wildland collections: 1) collect seed from a minimum of 50 randomly selected plants; 2) collect from widely separated individuals throughout a population without favoring the most robust or avoiding small stature plants; and 3) collect from all microsites including habitat edges (Basey et al. 2015). General collecting recommendations and guidelines are provided in online manuals (e.g., ENSCONET 2009; USDI BLM SOS 2021).
It is critical that wildland seed collection does not impact the sustainability of native plant populations. Collectors should take no more than 20% of the viable seed available at the time of harvest (USDI BLM SOS 2021). Additionally, care must be taken to avoid the inadvertent collection of weedy species, particularly those that produce seeds similar in shape and size to those of largeflower hawksbeard.
Post-Collection Management
Any vegetative material (e.g., stems, large leaves) should be removed from collected seed to reduce the moisture content and thus the potential for molding. This is especially important for largeflower hawksbeard because of the milky sap in vegetative material. Keep collections in dry, shaded locations, and do not allow seed to overheat. Seed should be loosely packed in collection bags to provide good air circulation. Seed can be dried by laying it out in thin layers on screens in a well-ventilated, rodent-free area. Potential seed insects should be controlled using appropriate chemicals or by freezing the seed collection for 48 hours once it is thoroughly dried. Temporary storage prior to delivery to the seed extractory should be in a cool (< 50 °F [10 °C]), dry (< 35% relative humidity) location.
Seed Cleaning
The following procedure was used by the Bend Seed Extractory to clean a very small seed lot (0.04 lb [18 g]) of largeflower hawksbeard (Barner 2009). Seed was first processed using a Westrup Model LA-P Laboratory Clipper/Deawner (Hoffman Manufacturing Corvallis, OR), which removed the seed from the seedheads. Seed was then air-screened using an office Clipper with a 1/14 × 1/2 top screen, 1/18 round bottom screen, medium speed, and medium air. The seed lot was finished using the Oregon Seed Blower (Hoffman Manufacturing Corvallis, OR) air column separator to remove the remaining nonviable seed and inert material. This procedure would likely be modified for a larger seed lot (Barner 2009).

Figure 10. Mature largeflower hawksbeard seed with scale (in cm). Photo: BLM OR030, SOS.
Seed Storage
Largeflower hawksbeard seed is orthodox. Dry seed stored for 2 months at (-4 °F [-20 °C) and 15% relative humidity had 89% viability (SER, INSR, RBGK 2023). Bend Seed Extractory stored largeflower hawksbeard at 33 to 38 °F (0.5-3 °C) (Barner 2009), but viability retention was not evaluated.
Seed Testing
A tetrazolium chloride (TZ) viability testing procedure was developed by the International Seed Testing Association for the hawksbeard genus (Moore 1985). There is no Association of Official Seed Analysts (AOSA) rule for testing germination.
Germination Biology
Germination studies were limited for largeflower hawksbeard. Maguire and Overland (1959) reported just 8% germination of untreated wildland seed. Seed was dried, threshed by hand, and stored in glass jars prior to germination testing that occurred within a year of harvest. Seed was put on moist blotter paper in petri dishes. Germination was 0% in dark conditions at 68 to 86 °F (20-30 °C). Germination was 8% in alternating light and dark conditions at 59 °F (15 °C), 8% for seed in dark at 68 °F (20 °C), and 8% for seed in alternating light-dark conditions at 68 to 86 °F (20-30 °C) (Maguire and Overland 1959).
Cold stratification improved the germination of at least some seed lots of tapertip hawksbeard (Crepis acuminata) and limestone hawksbeard (C. intermedia).
Wildland Seed Yield And Quality
Post-cleaning seed yield and quality of seed lots collected in the Intermountain region are provided in Table 2 (USDA FS BSE 2017). The results indicate that largeflower hawksbeard seed can generally be cleaned to high levels of purity, but seed fill and viability of fresh seed can be variable.
Table 2. Seed yield and quality of largeflower hawksbeard seed lots collected in the Intermountain region, cleaned by the Bend Seed Extractory, and tested by the Oregon State Seed Laboratory or the USDA Forest Service National Seed Laboratory (USDA FS BSE 2017).
| Seed lot characteristic | Mean | Range | Samples (no.) |
| Bulk weight (lbs) | 0.61 | 0.04-4.1 | 28 |
| Clean weight (lbs) | 0.12 | 0.005-0.5 | 28 |
| Clean-out ratio | 0.26 | 0.009-0.6 | 28 |
| Purity (%) | 99 | 93-99 | 28 |
| Fill (%)¹ | 88 | 70-99 | 28 |
| Viability (%)² | 92 | 75-98 | 26 |
| Seeds/lb | 112,997 | 83,300-152,000 | 28 |
| Pure live seeds/lb | 96,200 | 72,721-131,085 | 26 |
¹ 100 seed X-ray test
² Tetrazolium chloride test
Marketing Standards
Acceptable seed purity, viability, and germination specifications vary with revegetation plans. Purity needs are highest for precision seeding equipment used in nurseries, while some rangeland seeding equipment handles less clean seed quite well.
Agricultural Seed Production
Although a very small seed lot was harvested from seed production plots grown at USDA Forest Service Lucky Peak Nursery (Barner 2009), details about growing largeflower hawksbeard for agricultural seed production were lacking in the literature.
Hawksbeard species require 3 to 5 years to produce their first seed crop (Jensen et al. 2011).
Sears and Whitton (2016) reported that difficulties in producing nursery stock begin at the seed germination and establishment phases. Seedlings grow slowly, are sensitive to excess moisture and poor drainage, and produce long taproots making production of nursery stock and transplanting difficult (Babcock and Stebbins 1938).
Agricultural Seed Certification
In order to minimize genetic changes in specific accessions of native species when increased in cultivated fields, it is essential to track the geographic source and prevent inadvertent hybridization or selection pressure. This is accomplished by following third party seed certification protocols for Pre-Variety Germplasm (PVG) as established by the Association of Official Seed Certification Agencies (AOSCA). AOSCA members in the U.S., Canada, and other countries administer PVG requirements and standards that track the source and generation of planting stock. Field and cleaning facility inspections then monitor stand establishment, proper isolation distances, control of prohibited weeds, seed harvesting, cleaning, sampling, testing, and labeling for commercial sales (UCIA 2015; Young et al. 2020).
Seed growers apply for certification of their production fields prior to planting and plant only certified stock seed of an allowed generation (usually less than four). The systematic and sequential tracking through the certification process requires preplanning, knowing state regulations and deadlines, and is most smoothly navigated by working closely with state certification agency personnel. See the Wildland Seed Certification section for more information on stock seed sourcing.
Nursery Practice
The following procedure was used to grow largeflower hawksbeard seedlings for use in wildland restoration (Wirth 2000; Wirth and Pyke 2003). Seed was wild collected in south-central Oregon. It was cold stratified in moist sand (0.4 in [1 cm] deep) kept at 36 °F (2.2 °C) for 6 weeks, then moved to a greenhouse (68 °F [20 °C] day/night) to germinate. At cotyledon emergence, germinants were transplanted into Styrofoam vent blocks (0.75 in [1.9 cm] diameter, 5 in [12.7 cm] deep) filled with a mix of 30% potting soil and 70% loam. Transplants were grown for 10 weeks in the greenhouse, then moved to a cold frame to harden for 3 weeks. Seedlings were outplanted in late April and field survival was low. See the Wildland Seeding and Planting section for more information (Wirth 2000; Wirth and Pyke 2003).
Wildland Seeding And Planting
Largeflower hawksbeard is recommended for revegetation where precipitation averages 12 to 18 in (30-46 cm). The recommended seeding rate is 10 lb/ac (11 kg/ha) (Tilley et al. 2013).
Field studies conducted at Hart Mountain National Antelope Refuge, south-central Oregon, suggest that burned sites provided more favorable conditions for largeflower hawksbeard establishment than unburned sites (Wirth 2000; Wirth and Pyke 2003). The study area was on clayey soils and dominated by Wyoming big sagebrush/bluebunch wheatgrass vegetation. The study compared establishment and survival of seeded and transplanted largeflower hawksbeard at two different microsites on burned and unburned plots. Microsites included mounds beneath sagebrush canopies and interspaces between sagebrush canopies. Prescribed fires burned in late September 1997 and consumed 60 to 93% of understory vegetation.
Largeflower hawksbeard and other forb species (Modoc hawksbeard [C. modocensis] and woollypod milkvetch [Astragalus purshii]) were seeded or transplanted as seedlings grown from seed collected within 6 mi (10 km) of the study area (see Nursery Practice section for the procedure used to grow transplants) (Wirth 2000; Wirth and Pyke 2003). There were 250 seeds of each species seeded 0.4 in (1 cm) deep on each of the 8 sites (4 burned, 4 unburned). There were 200 seedlings planted at 3 burned and 3 unburned sites. Seedling emergence and survival and transplant survival were tracked through July 1999. The first growing season (1998) was longer with above average precipitation, and the second growing season (1999) was shorter with a cold spring followed by low precipitation. Crepis (largeflower and Modoc hawksbeard) emergence did not differ by species regardless of fire or microsite treatments (P = 0.87), so for seeding treatments Crepis species were analyzed together.
On unburned sites, Crepis emergence was significantly greater on mounds than in interspaces (P < 0.01) but this was followed by high mortality and lower overall establishment on unburned mounds (Table 3). Survival of seeded Crepis was greater on burned than unburned sites (P < 0.05) and greater on mounds than interspaces on burned and unburned sites (P < 0.1). Survival of seeded Crepis at the end of the second growing season was greatest (a little more than 70%) on burned mounds, next greatest (~65%) on burned interspaces, lower (~45%) on unburned mounds, and lowest (~35%) on unburned interspaces. Survival of transplanted largeflower hawksbeard seedlings was low (<14%), but better on burned than unburned sites (P < 0.1) and on mounds than interspaces (P < 0.05). Just seven Crepis plants flowered, and all of these were on burned sites (six on mounds and one in interspace) (Wirth 2000; Wirth and Pyke 2003).
Table 3. Mean emergence and survival from Crepis (largeflower and Modoc hawksbeard) seed and transplants at end of the second growing season on burned, unburned, mounds, and interspaces at Hart Mountain National Antelope Refuge, Oregon (Wirth 2000).
| Date/Cohort | Burned mounds | Burned interspaces | Unburned mounds | Unburned interspaces |
| ———————————-%———————————– | ||||
| Fall 1997 emergence | 40.7 | 33.3 | 50.2 | 33.6 |
| Fall 1997 seedling survival | 62.6 | 53.6 | 34.3 | 24.9 |
| Spring 1998 transplant survival | 22.4 | 15.2 | 9.7 | 8.1 |
Acknowledgements
Funding for Western Forbs: Biology, Ecology, and Use in Restoration was provided by the USDI BLM Great Basin Native Plant Materials Ecoregional Program through the Great Basin Fire Science Exchange. Great thanks to the chapter reviewers: Elizabeth R. Milano, Research Biological Scientist, USDA Forest Service, Rocky Mountain Research Station, and Stella Copeland, Research Ecologist, USDA Agricultural Research Service.
This research was supported in part by the USDA Forest Service, Rocky Mountain Research Station. The findings and conclusions in this publication are those of the authors and should not be construed to represent any official USDA or U.S. Government determination or policy.
Literature Cited
Abella, S.R. 2009. Smoke-cued emergence in plant species of ponderosa pine forests. Fire Ecology. 5(1): 22-37.
Andersen, B.A.; Holmgren, A.H. 1996. Mountain plants of northeastern Utah, revised. HG 506. Logan, UT: Utah State University Extension. 132 p.
Babcock, E.B.; Stebbins, G.L. 1938. The American species of Crepis: Their interrelationships and distribution as affected by polyploidy and apomixis. Pub. No. 504. Washington, DC: Carnegie Institution of Washington. 199 p.
Baker, W.L. 1983. Some aspects of the presettlement vegetation of the Piceance Basin, Colorado. The Great Basin Naturalist. 43(4): 687-699.
Baker, W.L.; Kennedy, S.C. 1985. Presettlement vegetation of part of northwestern Moffat County, Colorado, described from remnants. The Great Basin Naturalist. 45(4): 747-777.
Barner, J. 2009. Propagation protocol for production of propagules (seeds) Crepis occidentalis Nutt. Native Plant Network. U.S. Department of Agriculture, Forest Service, National Center for Reforestation, Nurseries, and Genetic Resources. http://npn.rngr.net/propagation/protocols [Accessed 2023 August 7].
Basey, A.C.; Fant, J.B.; Kramer, A.T. 2015. Producing native plant materials for restoration: 10 rules to collect and maintain genetic diversity. Native Plants Journal. 16(1): 37-53.
Bates, J.D.; Davies, K.W.; Sharp, R.N. 2007. Response of Wyoming big sagebrush communities to wildfire. In: Davies, K.W.; Nafus, A.M., eds. Sagebrush steppe – Research progress report 2007. Burns, OR: U.S. Department of Agriculture, Agricultural Research Service: 10-21.
Bates, J.D.; Rhodes, E.C.; Davies, K. 2011. Impacts of fire on sage-grouse habitat and diet resources. Natural Resources and Environmental Issues. 17(5): 1-17.
Bates, J.D.; Svejcar, T.J. 2009. Herbaceous succession after burning of cut western juniper trees. Western North American Naturalist. 69(1): 9-25.
Beatley, J.C. 1976. Vascular plants of the Nevada Test Site and central-southern Nevada: Ecologic and geographic distributions. TID-26881. Washington, DC: Technical Information Center, Energy Research and Development Administration. 306 p.
Blackwell, L.R. 2006. Great Basin wildflowers: A guide to common wildflowers of the high deserts of Nevada, Utah, and Oregon. Helena, MT: Morris Book Publishing. 288 p.
Bogler, D.J. 2006. Crepis. In: Flora of North America Editorial Committee, ed. Flora of North America North of Mexico. Volume 19 Magnoliophyta: Asteridae, part 6: Asteraceae, part 1. New York, NY: Oxford University Press: 214-228.
Bower, A.D.; St. Clair, J.B.; Erickson, V. 2014. Generalized provisional seed zones for native plants. Ecological Applications. 24(5): 913-919.
Charboneau, J.L.M.; Nelson, B.E.; Hartman, R.L. 2013. A floristic inventory of Phillips and Valley counties, Montana (U.S.A.). Journal of the Botanical Research Institute of Texas. 7(2): 847-878.
Consortium of Pacific Northwest Herbaria [CPNWH]. 2020. Seattle, WA: University of Washington Herbarium, Burke Museum of Natural History and Culture. pnwherbaria.org/
Coville, F.V. 1896. Crepis occidentalis and its allies. Contributions from the United States National Herbarium. 3(9): 559-565.
Cronquist, A. 1994. Volume five: Asterales. In: Cronquist, A.; Holmgren, A.H.; Holmgren, N.H.; Reveal, J.L.; Holmgren, P.K., eds. Intermountain flora: Vascular plants of the Intermountain West, U.S.A. Bronx, NY: The New York Botanic Garden. 496 p.
Davies, K.W.; Bates, J.D.; Miller, R.F. 2007. Environmental and vegetation relationships of the Artemisia tridentata spp. wyomingensis alliance. Journal of Arid Environments. 70(3): 478-494.
Davis, J.N. 1976. Ecological investigations in Cercocarpus ledifolius Nutt. communities of Utah. Brigham Young University. Thesis. 56 p.
Eldredge, E.; Novak-Echenique, P.; Heater, T.; Mulder, A.; Jasmine, J. 2013. Plants for pollinator habitat in Nevada. Tech. Note NV 57. Reno, NV: U.S. Department of Agriculture, Natural Resources Conservation Service. 65 p.
European Native Seed Conservation Network [ENSCONET]. 2009. ENSCONET seed collecting manual for wild species. Edition 1: 32 p.
Fertig, W.F.; Massatti, R.T.; Nelson, B.E.; Hartman, R.L. 2013. Annotated checklist of the vascular flora of the Wind River Range, Wyoming (U.S.A.). Journal of the Botanical Research Institute of Texas. 7(2): 905-939.
Gamble, L.H. 2008. Subsistence and feasting. In: Gamble, L.H., ed. The Chumash world at European contact: Power, trade, and feasting among complex hunter-gatherers. Berkeley, CA: University of California Press: 151-190.
Geary, A.B.; Merrill, E.H.; Cook, J.G.; Cook, R.C.; Irwin, L.L. 2017. Elk nutritional resources: Herbicides, herbivory, and forest succession at Mount St. Helens. Forest Ecology and Management. 401: 242-254.
Gregg, M.A.; Barnett, J.K.; Crawford, J.A. 2008. Temporal variation in diet and nutrition of preincubating greater sage-grouse. Rangeland Ecology and Management. 61(5): 535-542.
Hermann, F. 1966. Notes on western range forbs: Cruciferae through Compositae. Agric. Handb. 293. Washington, DC: U.S. Department of Agriculture, Forest Service. 365 p.
Hickman, J.C., ed. 1993. The Jepson manual: Higher plants of California. Berkeley, CA: University of California Press. 1400 p.
Hitchcock, C.L.; Cronquist, A. 2018. Flora of the Pacific Northwest: An illustrated manual. Second Ed. Giblin, D.E.; Legler, B.S.; Zika, P.F.; Olmstead, R.G., eds. Seattle, WA: University of Washington Press. 882 p.
James, J.J.; Drenovsky, R.E. 2007. A basis for relative growth rate differences between native and invasive forb seedlings. Rangeland Ecology and Management. 60(4): 395-400.
Jensen, S.; Stanton, S.; Nielson, M.; Stettler, J.; Mathews, D.; Ersfeld, T.J.; Sample, C. 2011. Selecting and growing Great Basin natives. In: Shaw, N.; Pellant, M., eds. Great Basin Native Plant Selection and Increase Project 2010 Progress Report. Boise, ID: U.S. Department of Agriculture, U.S. Forest Service, Rocky Mountain Research Station: 64-68.
Johnson, C.G.; Swanson, D.K. 2005. Bunchgrass plant communities of the Blue and Ochoco Mountains: A guide for managers. R6-NR-ECOL-TP-06-98. Portland, OR: U.S. Department of Agriculture, Forest Service, Pacific Northwest Research Station. 128 p.
Kelly, I.T. 1932. Ethnography of the Surprise Valley Paiute. University of California Publications in American Archaeology and Ethnology. 31(3): 67-210.
Kunzler, L.M.; Harper, K.T.; Kunzler, D.B. 1981. Compositional similarity within the oakbrush type in central and northern Utah. The Great Basin Naturalist. 41(1): 147-153.
Lady Bird Johnson Wildflower Center [LBJWC]. 2022. Crepis occidentalis Nutt. Native Plant Database. Austin, TX: Lady Bird Johnson Wildflower Center. https://www.wildflower.org/plants-main [Accessed 2023 August 7].
Lesica, P. 2012. Manual of Montana vascular plants. Fort Worth, TX: Botanical Research Institute of Texas (BRIT) Press. 779 p.
Luna, T.; Mousseaux, M.R.; Dumroese, R.K. 2018. Common native forbs of the northern Great Basin important for greater sage-grouse. Gen. Tech. Rep. RMRS-GTR-387. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station; Portland, OR: U.S. Department of the Interior, Bureau of Land Management, Oregon-Washington Region. 76 p.
Mackie, R.J. 1970. Range ecology and relations of mule deer, elk, and cattle in the Missouri River Breaks, Montana. Wildlife Monographs. 20: 3-79.
Maguire, A.D.; Overland, A. 1959. Laboratory germination of seeds of weedy and native plants. Washington State Agricultural Experiment Station Circular 349. Pullman, WA: Washington State University. 15 p.
Miller, R.F.; Krueger, W.C.; Vavra, M. 1981. Deer and elk use on foothill rangelands in northeastern Oregon. Journal of Range Management. 34(3): 201-204.
Miller, R.F.; Svejcar, T.J.; Rose, J.A. 2000. Impacts of western juniper on plant community composition and structure. Journal of Range Management. 53(6): 574-585.
Moore, R.P. 1985. Handbook on tetrazolium testing. Zurich, Switzerland: International Seed Testing Association. 99 p.
Munz, P.A.; Keck, D.D. 1973. A California flora and supplement. Berkeley, CA: University of California Press. 1905 p.
Murray, R.B.; Maryland, H.F.; Van Soest, P.J. 1978. Growth and nutritional value to cattle of grasses on cheatgrass range in southern Idaho. Ogden, UT: U.S. Department of Agriculture, Forest Service, Intermountain Forest and Range Experiment Station. 57 p.
Nowak, C.L.; Nowak, R.S.; Tausch, R.J.; Wigand, P.E. 1994. A 30000-year record of vegetation dynamics at a semi-arid locale in the Great Basin. Journal of Vegetation Science. 5(4): 579-590.
Omernik, J.M. 1987. Ecoregions of the conterminous United States. Map (scale 1:7,500,000). Annals of the Association of American Geographers. 77(1): 118-125.
Ott, J.; McArthur; E.D.; Roundy, B. 2003. Vegetation of chained and non-chained seedings after wildfire in Utah. Journal of Range Management. 56: 81-91.
Ott, J.E.; McArthur, E.D.; Sanderson, S.C. 2001. Plant community dynamics of burned and unburned sagebrush and pinyon-juniper vegetation in west-central Utah. In: McArthur, E. D.; Fairbanks, D. J., comps. Shrubland ecosystem genetics and biodiversity: Proceedings; 2000 June 13-15; Provo, UT. Proc. RMRS-P-21. Ogden, UT: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station: 177-191.
Passey, H.B.; Hugie, V.K. 1963. Some plant-soil relationships on an ungrazed range area of southeastern Idaho. Journal of Range Management. 16(3): 113-118.
Passey, H.B.; Hugie, V.K.; Williams, E.W.; Ball, D.E. 1982. Relationships between soil, plant community, and climate on rangelands of the Intermountain West. Tech. Bull. No. 1669. Washington, D.C. U.S. Department of Agriculture, Soil Conservation Service. 123 p.
Pearson, L.C. 1965. Primary production in grazed and ungrazed desert communities of eastern Idaho. Ecology. 46(3): 278-285.
Pennington, V.E.; Schlaepfer, D.R.; Beck, J.L.; Bradford, J.B.; Palmquist, K.A.; Lauenroth, W.K. 2016. Sagebrush, greater sage-grouse, and the occurrence and importance of forbs. Western North American Naturalist. 76(3): 298-312.
Plant Conservation Alliance [PCA]. 2015. National seed strategy for rehabilitation and restoration 2015-2020. Washington, DC: U.S. Department of the Interior, Bureau of Land Management. 52 p.
Pokorney, M.L.; Sheley, R.L.; Svejcar, T.J.; Engel, R.E. 2004. Plant species diversity in a grassland plant community: Evidence for forbs as a critical management consideration. Western North American Naturalist. 64(2): 219-230.
Reif, B.; Larson, J.; Jacobs, B.F.; Nelson, B.E.; Hartman, R.L. 2009. Floristic studies in north central New Mexico, U.S.A. the Tusas Mountains and Jemez Mountains. Journal of the Botanical Research Institute of Texas. 3(2): 921-961.
Richardson, B.; Kilkenny, F.; St. Clair, B.; Stevenson-Molnar, N. 2020. Climate Smart Restoration Tool. https://climaterestorationtool.org/csrt/
Rink, G.R.; Hodgson, W.; Phillips, B.G. 2020. Checklist of the vascular flora of the Kaibab Plateau, Coconino County, Arizona. Western North American Naturalist. 12(1): 1-52.
Sapsis, D.B. 1990. Ecological effects of spring and fall prescribed burning on basin big sagebrush/Idaho fescue-bluebunch wheatgrass communities. Corvallis, OR: Oregon State University. Thesis. 106 p.
Sears, C.J.; Whitton, J. 2016. A reexamination of the North American Crepis agamic complex and comparison with the findings of Babcock and Stebbins’ classic biosystematic monograph. American Journal of Botany. 103(7): 1289-1299.
SEINet – Regional Networks of North American Herbaria Steering Committee [SEINet]. 2020. SEINet Regional Networks of North American Herbaria. https://Symbiota.org/docs/seinet
Society for Ecological Restoration; International Network for Seed Based Restoration; Royal Botanic Gardens [SER, INSR, RBGK]. 2023. Seed Information Database (SID). Available from: https://ser-sid.org/
St. John, L.; Tilley, D. 2012. Plant guide: Limestone hawksbeard (Crepis intermedia) A. Gray. Aberdeen, ID: U.S. Department of Agriculture, Natural Resources Conservation Service, Aberdeen Plant Materials Center. 4 p.
Stebbins, G.L.; Jenkins, J.A. 1939. Aposporic development in the North American species of Crepis. Genetica. 21(3-4): 191-224.
Tilley, D.; Taliga, C.; Burns, C.; St. John, L. 2013. Plant materials for pollinators and other beneficial insects in eastern Utah and western Colorado. Tech. Note 2C. Boise, ID: U.S. Department of Agriculture, Natural Resources Conservation Service. 54 p.
USDA Forest Service, Bend Seed Extractory [USDA FS BSE]. 2017. Nursery Management Information System Version 4.1.11. Local Source Report 34-Source Received. Bend, OR: U.S. Department of Agriculture, Forest Service, Bend Seed Extractory.
USDA Forest Service, Western Wildland Environmental Threat Assessment Center [USFS WWETAC]. 2017. TRM Seed Zone Applications. Prineville, OR: U.S. Department of Agriculture, Forest Service, Western Wildland Environmental Threat Assessment Center. https://research.fs.usda.gov/pnw/products/dataandtools/seed-zone-webmap
USDA Natural Resources Conservation Service [USDA NRCS]. 2023. The PLANTS Database. Greensboro, NC: U.S. Department of Agriculture, Natural Resources Conservation Service, National Plant Data Team. https://plants.sc.egov.usda.gov/
USDI Bureau of Land Management, Seeds of Success [USDI BLM SOS]. 2020. Seeds of Success collection data. Washington, DC: U.S. Department of the Interior, Bureau of Land Management, Plant Conservation and Restoration Program.
USDI Bureau of Land Management, Seeds of Success [USDI BLM SOS]. 2023. Bureau of Land Management technical protocol for the collection, study, and conservation of seeds from native plant species for Seeds of Success. Washington, DC: U.S. Department of the Interior, Bureau of Land Management, Plant Conservation and Restoration Program. 45 p.
USDI Environmental Protection Agency [USDI EPA]. 2017. Ecoregions. Washington, DC: U.S. Department of the Interior, Environmental Protection Agency. https://www.epa.gov/eco-research/ecoregions
USDI Geological Survey [USDI USGS]. 2020. Biodiversity Information Serving Our Nation (BISON). U.S. Geological Survey. https://www.gbif.us/
Utah Crop Improvement Association [UCIA]. 2015. How to be a seed connoisseur. Logan, UT: UCIA, Utah Department of Agriculture and Food, Utah State University and Utah State Seed Laboratory. 16 p.
Welsh, S.L.; Atwood, N.D.; Goodrich, S.; Higgins, L.C., eds. 2015. A Utah Flora. Fifth Edition, revised. Provo, UT: Brigham Young University. 990 p.
Wentland, H.J. 1968. Summer range habits of the pronghorn antelope in central Montana with special reference to proposed sagebrush control study plots. Bozeman, MT: Montana State University. Thesis. 75 p.
Whitton, J.; Sears, C.J.; Maddison, W.P. 2017. Co-occurrence of related asexual, but not sexual, lineages suggests that reproductive interference limits coexistence. Proceedings: Biological Sciences. 284(1868): 1-8.
Wirth, T.A. 2000. Emergence, survival, and reproduction of three species of forbs important to sage grouse nutrition in response to fire, microsite, and method of establishment. Corvallis, OR: Oregon State University. Thesis. 62 p.
Young, S.A.; Schrumpf, B.; Bouck, M.; Moore, M. 2020. How the Association of Official Seed Certifying Agencies (AOSCA) tracks wildland sourced seed and other plant propagating materials. Logan, UT: Utah State University, Utah Agricultural Experiment Station. 36 p.
How to Cite
Gucker, Corey L.; Shaw, Nancy L. 2023. Largeflower hawksbeard (Crepis occidentalis). In: Gucker, C.L. ; Shaw, N.L., eds. Western forbs: Biology, ecology, and use in restoration. Reno, NV: Great Basin Fire Science Exchange. Online: https://westernforbs.org