Authorship
Gucker, Corey
Publication Date
January 2019
Nomenclature
Sphaeralcea coccinea (Nuttall) Rydberg is commonly referred to as scarlet globemallow. It belongs to the mallow or Malvaceae family (LaDuke 2016).
Family
Malvaceae – Mallow family
Genus
Sphaeralcea
Species
coccinea
NRCS Plant Code
SPCO (USDA NRCS 2017).
Subtaxa
The Flora of North America (LaDuke 2016) recognizes two scarlet globemallow varieties: coccinea and elata (Baker f.) Kearney.
Synonyms
Malva coccinea Nuttall, Malvastrum coccineum (Nuttall) A. Gray, Sida coccinea (Nuttall) de Candolle (LaDuke 2016).
S. coccinea var. coccinea: Malvastrum coccineum (Nuttall) A. Gray var. dissectum (Nuttall ex Torrey & A. Gray), M. cockerellii A. Nelson, M. dissectum (Nuttall ex Torrey & A. Gray) Cockerell var. cockerellii (A. Nelson) A. Nelson, Sida dissecta Nuttall ex Torrey & A. Gray, Sphaeralcea coccinea subsp. dissecta (Nuttall ex Torrey & A. Gray) Kearney, S. coccinea var. dissecta (Nuttall ex Torrey & A. Gray) Kearney.
S. coccinea var. elata: Malvastrum coccineum (Nuttall) A. Gray var. elatum Baker f., M. elatum (Baker f.) A. Nelson, M. micranthum Wooton & Standley, Sphaeralcea coccinea subsp. elata (Baker f.) Kearney, S. elata (Baker f.) Rydberg (LaDuke 2016).
Common Names
Scarlet globemallow, caliche globemallow, common globemallow, red false mallow, scarlet falsemallow (Craighead et al. 1963; Johnson and Nichols 1970; Spellenberg 2001; LaDuke 2016; LBJWC 2018).
Chromosome Number
Chromosome numbers are: 2n = 10, 20 (Holmgren et al. 2005), or 30 (Pendery and Rumbaugh 1993). Generally, variety coccinea is 2n = 10 and elata is 2n = 20 (LaDuke 2016).
Hybridization
Hybridization within the Sphaeralcea genus is common. Scarlet globemallow and gooseberryleaf globemallow (S. grossulariifolia) intermediates occur where distributions overlap (Atwood and Welsh 2002).
Distribution
Scarlet globemallow is the most widely distributed globemallow species (Sphaeralcea) in North America (Pendery and Rumbaugh 1993). It occurs from British Columbia to Manitoba and south to Arizona, New Mexico, Texas, and Chihuahua, Mexico (La Duke 2016). It is more common in the Great Plains than in the Intermountain region (Pendery and Rumbaugh 1993). Variety coccinea is found throughout the species range but elata occurs only in New Mexico, Texas, and northern Mexico (La Duke 2016).
Scarlet globemallow is considered more common on the eastern than the western side of the Rocky Mountains (Craighead et al. 1963), although in the state of South Dakota it becomes more abundant as one moves westward (Johnson and Nichols 1970) and in Utah, it occurs in all but Morgan County (Welsh et al. 1987).
Habitat And Plant Associations
Scarlet globemallow occurs in grassland, shrubland, and open woodland vegetation throughout its range. It is generally most abundant in open communities and disturbed sites within the 8- to 12-in (200–300 mm) precipitation zones (Wasser 1982) and frequently occurs along roadsides (Craighead et al. 1963; Johnson and Nichols 1970; La Duke 2016).
Scarlet globemallow is common in western and central US grasslands including desert grasslands dominated by black grama (Bouteloua eriopoda), shortgrass prairies with blue grama (B. gracilis) and buffalograss (Buchloe dactyloides), northern mixed-grass prairies dominated by needle and thread (Hesperostipa comata) and western wheatgrass (Pascopyrum smithii), central mixed-grass prairies with western wheatgrass, blue grama, and buffalograss, and southern mixed-grass prairies with little bluestem (Schizachyrium scoparius), big bluestem (Andropogon gerardii) and sideoats grama (B. curtipendula) (Sims et al. 1978; Shiflet 1994). In the Northern Rocky Mountains, scarlet globemallow is common in bluebunch wheatgrass (Pseudoroegneria spicata)-blue grama, bluebunch wheatgrass-western wheatgrass, and needle-and-thread-blue grama grasslands (Shiflet 1994). In the Great Plains of southern Alberta and Saskatchewan, scarlet globemallow is a principal forb in mixed-grass prairie types, where it is inconspicuous in climax prairie vegetation but profuse and forms large populations in disturbed areas (Coupland 1950).
Throughout its range, scarlet globemallow is also common in a variety of shrublands (Fig. 1) and woodlands. In the Intermountain West, scarlet globemallow occurs with greasewood (Sarcobatus spp.), saltbush (Atriplex spp.), sagebrush (Artemisia spp.), pinyon-juniper (Pinus–Juniperus spp.), and Gambel oak (Quercus gambelii) (Holmgren et al. 2005). In western Montana, it is common with bluebunch wheatgrass, western wheatgrass, and skunkbush sumac (Rhus trilobata) and often increases in abundance with grazing (Mueggler and Stewart 1980). It is also a noted forb in ponderosa pine (Pinus ponderosa)/bluebunch wheatgrass woodlands in the Little Belt Mountain foothills in Meagher County, Montana (Daubenmire 1952). In a survey of grasslands and shrublands in Wyoming and Colorado, it was important in abandoned agricultural lands, blue grama short-grass prairies, dry meadows occupying alkaline swales near streambanks, mixed grass-sedge, big sagebrush (Artemisia tridentata), pinyon-juniper, and browse shrubland types dominated by Gambel oak and alderleaf mountain mahogany (Cercocarpus montanus) (Costello 1944b). In Utah, blackbrush (Coleogyne ramosissima), salt desert brush, mixed desert shrub, pinyon-juniper, and ponderosa pine vegetation types are common scarlet globemallow habitat (Welsh et al. 1987).

Figure 1. Scarlet globemallow growing in typical shrubland/grassland habitat in Utah. Photo: USDI Bureau of Land Management (BLM) UT080 Seeds of Success (SOS).
Although generally considered intolerant of shade and restricted to woodland interspaces (Wasser 1982), scarlet globemallow occurs beneath tree canopies in parts of New Mexico and Arizona. In twoneedle pinyon (P. edulis) and one-seed juniper (J. monosperma) woodlands in south-central New Mexico, scarlet globemallow was associated with one-seed juniper. Basal cover of scarlet globemallow was 0% near the bole, 0.2% beneath the canopy, and 0.15% in the interspaces. Basal cover was significantly greater (P ≤ 0.05) beneath the tree canopy and in the interspaces than within 3.3 ft (1 m) of the tree boles. (Armentrout and Pieper 1988). When vegetation was compared beneath and outside of tree canopies in central Arizona’s Verde River Watershed, scarlet globemallow was more abundant beneath velvet mesquite (Prosopis velutina) canopies than outside of the canopies on the terraces between the desert scrub and riparian communities (Schade et al. 2003).
Elevation
Scarlet globemallow occupies sites from 980 to 9,020 ft (300–2,750 m) elevation (Holmgren et al. 2005; La Duke 2016). In Utah, the species is found at 3,020 to 9,020 ft (920–2,750 m) elevation (Welsh et al. 1987). Variety elata occurs from 980 to 5,250 ft (300–1,600 m) in New Mexico and Texas, and coccinea occurs across the elevation range of the species (La Duke 2016).
Soils
Scarlet globemallow grows in a broad range of soil types and textures but often occurs on dry, sandy, gravelly sites (Wasser 1982; Stubbendieck et al. 1986; Holmgren et al. 2005). It tolerates low to moderate salinity levels but not sodic or consistently waterlogged soils (Hladek et al. 1972; Wasser 1982). The more geographically restricted elata variety grows in rocky, sandy, and silty soils of limestone, gypsum, or igneous substrates (La Duke 2016).
Scarlet globemallow occupies soils ranging from sands to clays throughout its range (Hanson 1955; Hanson and Dahl 1957; Sims et al. 1978; Sieg et al. 1983). In a greenhouse study, however, short-term survival of scarlet globemallow was significantly greater (P = 0.009) in sandy loam (71% survival) than clay loam (32% survival) field-collected soils. Sandy loams were 69% sand, 13% clay, and 19% silt, and clay loams were 35% sand, 30% clay, and 36% silt (Rawlins et al. 2009).
Dry sites are common habitat for scarlet globemallow, which is considered drought resistant and most frequent at sites receiving 7 to 12 in (180–300 mm) of annual precipitation (USDA NRCS IDPMC 2005). Along a gradient of arid to dry to wet-mesic grasslands in Saskatchewan, scarlet globemallow was an indicator of dry conditions (Looman 1963). When vegetation was compared along a moisture gradient in 65 mixed-grass prairie stands in the shale-limestone region of north-central Kansas, basal cover of scarlet globemallow was greatest in very dry stands (5.9%). Cover was less in dry (3.6%), dry-mesic (1.1%), and mesic or wet-mesic (0%) stands. Dry stands had loam or silt loam soils with pH of 7.8 to 8. Sand and rock contents were greatest in the soils of very dry stands, which occurred on southern or western slopes of 10° to 29° (Hladek et al. 1972). In contrast, in mixed-grass prairies of northwestern Kansas and southwestern Nebraska, frequency of scarlet globemallow was 7.8% on very dry, 1.8% on dry, 7.7% on dry mesic, and 10.8% on mesic sites with loamy to very fine sandy loam soils (Hulett et al. 1968). When slope-plant relationships were evaluated in blue grama grasslands of North Dakota’s Little Missouri Badlands, scarlet globemallow frequency was greatest on the shallowest slopes. At this site soils were excessively drained loam and clay soils often less than 5 in (13 cm) deep. Frequency was 32% on west slopes of 3°, 15% on east slopes of 8°, and 5% on east and west slopes of 11° and 16° (Dix 1958). For the previous studies, site dryness was characterized by soil composition and slope, soil moisture was not directly measured.
Several studies report scarlet globemallow growing in somewhat saline soils. In the Great Plains of southern Alberta and Saskatchewan, scarlet globemallow abundance was greatest in mixed-grass prairies with clay loam, solonetzic soils. Sodium is characteristic in the upper horizons of solonetzic soils (Coupland 1950). In Carter County, Montana, scarlet globemallow was the most common forb in the sagebrush-saltbrush/southeastern grassland vegetation type on clay-clayey-saline sites (Sieg et al. 1983). In a study of vegetation and soil relationships in blue grama stands on the Central Plains Experimental Range near Nunn, Colorado, abundance of scarlet globemallow increased with decreasing soil permeability and grazing on saline-alkali soils. Frequency of scarlet globemallow, though, was greater in sandy loams or loams (29–43%) than in clay loam or saline-alkalis (9–12%) (Hyder et al. 1966). Soil salinity levels can be too high to support scarlet globemallow populations. One-year-old transplants did not survive four growing seasons on unamended bentonite mine spoils near Upton, Wyoming. At this site, soils were highly saline or sodic with an average pH of 6.9 (range: 4.1–8) (Uresk and Yamamoto 1994).
In an evaluation of vegetation and soil relationships in 89 stands across the central Great Plains, occurrence of scarlet globemallow was positively correlated (P < 0.05) with soil phosphorus. The species was most common in the Central High Plains of eastern Wyoming and eastern Colorado and Central High Tablelands of eastern Colorado, western Kansas, and southwestern Nebraska (Nicholson and Hulett 1969).
Description
Scarlet globemallow is a perennial with a slender, deep taproot, woody branched caudex, and sometimes well-developed rhizomes (Hermann 1966; La Duke and Northington 1978; Holmgren et al. 2005; La Duke 2016).
Aboveground. Plants produce few to many ascending or low, spreading, slender stems. Stems may reach 24 in (61 cm) but are typically less than 12 in (30 cm) tall (La Duke and Northington 1978; Spellenberg 2001; La Duke 2016; LBJWC 2018). When plants were monitored for 8 years at the Dickinson Experiment Station in southwestern North Dakota, scarlet globemallow plants reached a minimum height of 4 in (10 cm) and a maximum of 5 in (13 cm) in July (Goetz 1963). Stems, which are unbranched or less commonly short-branched, arise from the caudex or sometimes from rhizomes (Welsh et al. 1987; Holmgren et al. 2005; Pavek et al. 2012; La Duke 2016). Colony formation and extensive rhizome development have been described (Hermann 1966; Holmgren et al. 2005; LBJWC 2018).
Stems and leaves are generally covered in dense stellate hairs, which make plants appear light green to grayish (Hermann 1966; Welsh et al. 1987; La Duke 2016). Basal leaves are lacking, and stem leaves are alternate and petiolate (Welsh et al. 1987; Holmgren et al. 2005). Generally, leaf blades are slightly wider than they are long, but thickness, divisions, and pubescence varies among populations (La Duke and Northington 1978; Stubbendieck et al. 1986; Spellenberg 2001; Holmgren et al. 2005). Leaf blades can be 0.5-2 in (1.2–5.2 cm) wide and almost as a long with three to five narrow to broad lobes (see Figs. 2, 4, 6, 7) (Stubbendieck et al. 1986; Spellenberg 2001; Holmgren et al. 2005).
Flowers occur in narrow clusters in a terminal raceme or sometimes a paniculate inflorescence (Fig. 4) (Hermann 1966; Welsh et al. 1987; Spellenberg 2001). Lower flowers are generally solitary in the leaf axils, while upper flowers have short pedicels (2–10 mm) that develop in the inflorescence as it elongates up to 4 in (10 cm) (Hermann 1966; Stubbendieck et al. 1986; Holmgren et al. 2005). Individual flowers are perfect, radially symmetric, and about 1-in (2.5 cm) wide. Flowers have five deep orange to brick red petals (Stubbendieck et al. 1986; Spellenberg 2001; Holmgren et al. 2005) and numerous yellow stamens, which are joined at the base around the style (Spellenberg 2001; La Duke 2016). The staminal column is 3 to 5 mm long (Holmgren et al. 2005). Fruits are segmented schizocarps (4.8–6.7 mm in diameter) (Fig. 3), which split into 8 to 14 mericarp sections (Welsh et al. 1987; Holmgren et al. 2005; La Duke 2016). The schizocarps and their mericarps have stellate pubescence, similar to that of the stems and leaves (Craighead et al. 1963). Mericarps are prominently veiny and contain one kidney shaped, brown to black seed (Stubbendieck et al. 1986; Holmgren et al. 2005; La Duke 2016).

Figure 2. Scarlet globemallow growing in gravelly soil in Wyoming. Photo: BLM WY030 SOS.
Irregularities in plant growth related to site conditions and hybridization where species overlap make distinguishing Sphaeralcea species difficult, but some tips have been provided. Scarlet globemallow is often one of the first globemallows to flower (La Duke 2016). Production of a single flower per node is characteristic of scarlet globemallow and helpful in distinguishing it from related species (Sriladda et al. 2012). Also, scarlet globemallow mericarps have 66% of their faces covered by with rough ridges while other globemallows have 50% or less (Hermann 1966). Scarlet globemallow varieties can be distinguished by careful examination of the leaves. Variety coccinea produces mid- and secondary leaf lobes of nearly equal lengths, while mid-lobe leaf lengths exceed that of the secondary lobes for variety elata (La Duke 2016).

Figure 3. Scarlet globemallow schizocarps. Scale in mm. Photo: BLM WY 932A SOS.
Belowground. Scarlet globemallow produces slender, deep taproots with limited branching and lateral development. Water is likely absorbed along the entire length of the taproot because it is fleshy and shrinks rapidly with exposure to air (Weaver 1958). When eight plants were excavated from three prairies in southwestern Saskatchewan, roots were 2 to 5 mm in diameter at the soil surface and tapered to 1 mm at 35 to 47-inch (90-120 cm) depths. The maximum rooting depth was 5.9 feet (1.8 m) (Coupland and Johnson 1965). In short-grass prairie in west-central Kansas, scarlet globemallow roots were slender and unbranched until about 3 feet (1 m) deep where they divided into laterals spanning about 1-foot (0.3 m). Taproots reached 7.5 feet (2.3 m) deep (Albertson 1937).
Additional excavation studies suggest that root growth may vary with moisture availability and grazing. Plants excavated from short-grass prairie in the central Great Plains following an 8-year drought produced roots that reached 7 feet (2.1 m) but were more commonly 4 feet (1.2 m) deep (Weaver and Albertson 1943). When eight plants were excavated from ungrazed prairie in the same region, roots penetrated 8 to 13 feet (2.4-4 m) deep and had little branching or lateral spread (Weaver 1958). Plants excavated from heavily grazed mixed-grass prairie near Ashland and Atwood, Kansas, reached depths of just over 4 feet (1.2 m) (Tomanek and Albertson 1957).
Research indicates that scarlet globemallow roots are colonized by arbuscular mycorrhizal fungi (AMF). In experiments, 64% of scarlet globemallow roots were colonized with AMF after 120 days, but plant growth did not differ significantly in AMF- and AMF+ soils (Busby et al. 2011).
Reproduction
Scarlet globemallow reproduces by seed. Plants generally begin growing in March or April, produce flowers from May to July, and develop most seed in July and August throughout the scarlet globemallow range (St. John and Ogle 2009). A single large plant growing without competition in North Dakota produced 1,860 seeds when harvested at a time of likely maximum seed production (Stevens 1957). Once seedlings are established, however, plants can spread by rhizomes. In a disturbed site in the Piceance Basin in northwestern Colorado, researchers found young plants connected to parent plants by rhizomes extending 3.3 ft (1 m) laterally. In this same area, scarlet globemallow seed production was prolific with abundant seasonal moisture but declined with decreasing moisture availability (Grygiel et al. 1984).
Phenology
Although flowering is most common in late spring and early summer, flowering of scarlet globemallow can occur from March through October throughout its broad range (Craighead et al. 1963; Dickinson and Dodd 1976; Hewitt 1980; Kemp 1983; Stubbendieck et al. 1986; Spellenberg 2001; Holmgren et al. 2005; USU Ext. 2017; LBJWC 2018). Harvestable seed is often available in July and August (Belcher 1985; USDI BLM SOS 2017). Timing of flowering and seed production can vary with site conditions, elevation, and climate (Wasser 1982; Kemp 1983), but spring and summer flowering is most common for both varieties (Craighead et al. 1963; Wasser 1982; White 1995; La Duke 2016; USDI BLM SOS 2017).
Flowers occur from late April to October in the Intermountain West (Holmgren et al. 2005) and from May to early July in the Rocky Mountains (Craighead et al. 1963). In the Great Plains of southern Alberta and Saskatchewan, scarlet globemallow flowering starts in early June and lasts about a month (Coupland 1950). The average flowering date for scarlet globemallow was June 7 based on 4 years of monitoring at Swift Current, Saskatchewan, and June 16 based on 13 years of monitoring at Treesbank, Manitoba (White 1995). In 2 years of monitoring of mixed-grass prairie in south-central Montana, scarlet globemallow flowering began between May 20 and June 8 and lasted 20 to more than 27 days (Hewitt 1980). Over 3 or 4 years of monitoring in Woodworth, North Dakota, the earliest first bloom date was June 7 and the latest was July 7. Median date at full flower was June 15, and flowering was 95% complete on July 10. The flowering period averaged 25 days (Callow et al. 1992).
Breeding System
Flowers are strongly outcrossing (Fig. 4), and bees, especially Diadasia species, are important pollinators. In greenhouse experiments, scarlet globemallow produced few schizocarps and little seed when self-pollinated (Cane 2009). When outcrossed, flowers produced 8 to 10 times as many schizocarps and five to nine times as much seed as protected flowers. Indeterminate flowering and a broad elevational and geographical range suggest that scarlet globemallow supports a diversity of pollinators. In wild populations, a diversity of bees visit scarlet globemallow flowers. Two sunflower bee (Diadasia spp.) specialists occurred in every globemallow patch visited in Nevada, eastern Oregon, southern Idaho, and northwestern Utah (Cane 2009). Scarlet globemallow flower nectar is also a food source for mature small-checkered skipper butterflies (Pyrgus scriptura) (LBJWC 2018).

Figure 4. Scarlet globemallow inflorescences and flowers. Photo: BLM WY020 SOS.
Ecology
Scarlet globemallow is drought and disturbance tolerant. It typically increases in abundance on disturbed sites yet persists in climax communities (Albertson 1937). Scarlet globemallow was considered an aggressive invader of disturbed sites in the Piceance Basin, northwestern Colorado (Grygiel et al. 1984). Three years following disturbances designed to mimic energy development, scarlet globemallow seedlings and relatively small, mature plants occurred as relatively large clumps or islands with multiple plants. In the next 2 years, island size decreased. Researchers concluded that scarlet globemallow aggressively invaded open sites but did not aggressively compete with established plants. Though abundance decreased with competition from mature neighboring plants, scarlet globemallow remained a component of the vegetation (Grygiel et al. 1984). In a study of 26 mixed-grass prairies ranging from early seral to near climax, production of scarlet globemallow was greatest in early seral (81 lbs/ac [91 kg/ha]) communities but remained at about half of that throughout mid-seral (40–42 lbs/ac [45–47 kg/ha]) and climax (39 lbs/ac [44 kg/ha]) conditions (Uresk 1990).
Taproots are the primary carbohydrate storage organ, and plants generally survive defoliation events and shed leaves during dry periods (Menke and Trlica 1981; Stubbendieck et al. 1986). When evaluated for 2 years in shortgrass and antelope bitterbrush (Purshia tridentata)-sagebrush exclosures in northern Colorado, the mass of scarlet globemallow roots and rhizomes was many times that of crowns. With stem elongation in the spring, total non-structural carbohydrate (TNC) levels measured a low of 58 mg/g. Drawdowns in TNC also occurred during fruit development and seed shatter stages. High TNC levels (171 mg/g) were reached in November, and TNC replenishment was rapid. An increase from 70 mg/g in early September to 171 mg/g in mid-November was measured in one year (Menke and Trlica 1981). Herbage yield, plant vigor, and TNC levels of defoliated scarlet globemallow plants (90% removed) were not significantly different from control plants by 14 months after defoliation (Trlica et al. 1977).
Scarlet globemallow is referred to as a long-lived species (Kitchen 1994; St. John and Ogle 2009), and in modelling studies, a maximum age of 35 was used for analysis of scarlet globemallow populations (Coffin and Urban 1993). Based on long-term mapping of permanent plots in mixed-grass prairie near Hays, Kansas, however, researchers suggested the maximum life span of scarlet globemallow was 6 years. Survival from year 1 to year 2 was 28% based on observations of 290 plants in livestock exclosures (Lauenroth and Adler 2008).
Disturbance Ecology
Many studies reveal at least short-term increases in scarlet globemallow abundance following disturbances including droughts, burning, grazing, and other soil disturbances (Albertson et al. 1957). Increases in scarlet globemallow abundance often coincide with decreases in associated vegetation (Albertson and Weaver 1944).
Scarlet globemallow abundance increased with drought conditions and associated grass losses at several mixed- and short-grass prairies in the central Great Plains. In long-term studies that included drought (1933–1940) and mesic (1941–1944) conditions, scarlet globemallow abundance increased with drought conditions, which were associated with dramatic decreases in grass abundance. Early spring growth, loss or shriveling of leaves in the absence of moisture, and regeneration from surviving underground tissue allowed scarlet globemallow to survive severe and long-lasting drought conditions (Albertson and Weaver 1944). On the Central Plains Experimental Range near Nunn, Colorado, scarlet globemallow dominated patches where blue gramma plants were senescing or dead in grazed and ungrazed short-grass prairie (Stanton et al. 1984). In the same area, density of scarlet globemallow was four times greater for 6 years on sites with all blue grama tillers removed than on sites without removal (P < 0.05) (Munson and Lauenroth 2009).
Dormant-season fires generally result in increases or unchanged scarlet globemallow populations, but growing-season fires may reduce abundance. After an August fire in dry mixed-prairie in southeastern Alberta, scarlet globemallow abundance was negligibly different on burned and unburned sites (Brockway et al. 2002). For the first 3 post-fire years, abundance was 0.3 to 0.6% greater on burned than unburned sites, which were grazed by cattle before and after burning (0.64 AUM/ha) (Erichsen-Arychuk et al. 2002). In badland grasslands of western North Dakota, frequency of scarlet globemallow increased following a late September fire and was relatively unchanged following a May fire. Three years after the September fire, scarlet globemallow frequency was 42% on burned and 12% on unburned plots with low to moderate grazing pressure. Three months after the May fire, frequency was 12% on burned and 10% on unburned plots with light grazing pressure (Dix 1960). Cover increased followed a dormant-season burn (1.2%) and decreased followed a growing-season burn (0.3%) as compared to unburned plots (0.8%) in short-grass prairie at the Kiowa National Grassland, northeastern New Mexico. Differences between post-fire effects were evaluated 3 to 5 months after the fire and were significantly different from controls (P ≤ 0.05) (Brockway et al. 2002). On the Custer Battlefield National Monument, south-central Montana, frequency of scarlet globemallow was 15 to 18% on burned and 5 to 8% on nearby unburned plots following the August fire. Burned and unburned differences were significant for all 3 post-fire years (P < 0.05) (Bock et al. 1986).
Scarlet globemallow is often an early colonizer of abandoned agricultural sites or other soil disturbances. In western North Dakota agricultural fields abandoned 1 to 35 years, scarlet globemallow was an important old-field species. It was usually present from the earliest stages of recovery to climax short-grass vegetation dominance (Whitman et al. 1943). In eastern Wyoming and eastern Colorado, scarlet globemallow was common on cultivated sites within 6 to 10 years of abandonment. In a chronosequence, frequency was 19% about 5 years after abandonment and increased to 60 to 90% from 9 to 40 years after abandonment (Costello 1944a,b). At Wyoming’s High Plains Grasslands Research Station, scarlet globemallow occurred on all mixed-grass prairie sites disturbed 1 to 51 years earlier. Frequency was 4.6% on 1-year-old and 7.8% on 51-year-old disturbed sites, which were either scraped or cultivated (Samuel and Hart 1994).
Scarlet globemallow survival or colonization was greater on less severely disturbed soils when progressively more aggressive soil manipulations were compared in a big sagebrush-grassland in the Piceance Basin, Colorado. Vegetation recovery was monitored for 6 years following four treatments: 1) soil as undisturbed as possible following vegetation removal; 2) soil scarified to 12 in (30 cm) deep; 3) soil to 3 ft (1 m) removed, mixed, and replaced; 4) soil to 7 ft (2 m) removed, inverted, and replaced (Table 1; Biondini et al. 1985).
Table 1. Canopy cover (%) of scarlet globemallow in a big sagebrush-grassland in the Piceance Basin, CO, as time since progressively more aggressive soil treatments increased (Trt 1- least soil disturbance – Trt 4 most soil disturbance) (Biondini et al. 1985).
|
Time since treatment (yrs) |
1 |
4 |
5 |
6 |
|
———Cover %——– |
||||
|
Veg removed, little soil disturbance |
27 |
28 |
22 |
23 |
|
Soil scarified to 30 cm |
12 |
18 |
33 |
10 |
|
Soil to 1 m removed, mixed, and replaced |
1 |
2 |
4 |
4 |
|
Soil to 2 m removed, inverted, and replaced |
1 |
2 |
4 |
3 |
Wildlife And Livestock Use
Scarlet globemallow is an important food source for a variety of wildlife and livestock. It is considered excellent forage for deer (Odocoileus spp.) and pronghorn (Antilocapra americana) and fair forage for livestock. Importance to livestock is generally greater in the Southwest than in the Great Plains, but increases occur in both areas with drought and heavy grazing (Johnson and Nichols 1970; Stubbendieck et al. 1986; Pendery and Rumbaugh 1993). Scarlet globemallow seeds are eaten by many birds and small mammals (LBJWC 2018).
Ungulates. Scarlet globemallow is palatable and important to pronghorn even when it is not a dominant vegetation component (Buechner 1950; Stephenson et al. 1985). At the Pawnee National Grassland in northeastern Colorado, relative density of scarlet globemallow in collected pronghorn fecal samples was 2% in May, 32% in June, 37% in August, and 58% in October. Scarlet globemallow was not abundant in the study area (Schwartz and Nagy 1976). Another study in the Pawnee National Grassland, reported scarlet globemallow as the largest single dietary occurrence in pronghorn diets (18% in summer) (Ellis and Travis 1975). In cattle-grazed short-grass prairie in western Kansas, high amounts of scarlet globemallow occurred in pronghorn fecal samples collected in May (20%) and October (21%) (Sexson et al. 1981). In Oldham and Hartley counties in the Texas Panhandle, scarlet globemallow was the most important forb in pronghorn diets. Scarlet globemallow made up 12% of spring, 15% of summer, 12% of fall, and 3% of winter diets for 1 year (Koerth et al. 1984). In desert scrub and grasslands of south-central New Mexico, scarlet globemallow was important in pronghorn and non-native African onyx (Onyx gazella) diets (Cain et al. 2017). In north-central New Mexico, scarlet globemallow was 6% of the composition of pronghorn diets averaged over 2 years and all seasons. Scarlet globemallow cover was 1% and frequency 4% in the study area (Stephenson et al. 1985).
Deer and elk (Cervus canadensis) use of scarlet globemallow was reported at several sites in New Mexico (Mahgoub 1984; Sandoval et al. 2005; Bender et al. 2007), and bison (Bos bison) utilization was reported in Colorado (Peden 1976). Scarlet globemallow is considered a preferred forb of mule deer (O. hemionus) in New Mexico (Bender et al. 2007). Scarlet globemallow was important in mule deer diets in New Mexico’s Sacramento Mountains where it was less than 1% of the vegetation. From fecal samples collected from June 1981 through September 1983, scarlet globemallow was 2.4% of annual diets (Mahgoub 1984). Although scarlet globemallow occurred with very little cover in ponderosa pine and twoneedle pinyon-oneseed juniper woodlands in north-central New Mexico, it comprised 4% of elk and 6% of mule deer annual diets (Sandoval et al. 2005). In short-grass prairie at the Pawnee National Grassland, scarlet globemallow was consumed most by bison in June (5.6–6.7% of diets) on lightly and heavily grazed sites evaluated from December through October (Peden 1976).
Small mammals. Black-tailed prairie dogs (Cynomys ludovicianus), desert cottontails (Sylvilagus audubonii), black-tailed jackrabbits (Lepus californicus), and a variety of rodents feed on scarlet globemallow. It is particularly important to prairie dogs in the summer. At two short-grass prairie sites in central Colorado, scarlet globemallow was the most abundant forb in prairie dog diets. It was consumed in all 4 months (May, June, August, and September) of the study, and at one site, made up 24% of the relative density of September diets (Bonham and Lerwick 1976). Scarlet globemallow was also a major part of prairie dog diets between March and September in short-grass prairie in South Dakota’s Conata Basin. The percent composition of scarlet globemallow in fecal samples averaged 10% from March through September (range: 3-21%) where it was a dominant forb making up 77% frequency and 6% cover of the vegetation (Uresk 1984). In two prairie dog towns in western South Dakota, occurrence of scarlet globemallow in prairie dog stomachs often exceeded its occurrence in the study area. Scarlet globemallow made up 11 to 33% of the dry weight of stomach samples taken in December, May, June, July, and August (Summers and Linder 1978). In Buffalo Gap National Grassland in South Dakota, the abundance of scarlet globemallow in prairie dog diets was significantly greater in September (21%) than any other month (Dec-Feb, May, and July) when it was 0.6 to 2.8% of diets (Fagerstone et al. 1981).
Several studies report substantial use of scarlet globemallow by desert cottontails and often lower levels of use by black-tailed jackrabbits. Scarlet globemallow made up a large percentage of desert cottontail diets when stomach contents were analyzed from collections made in mountain mahogany and short-grass prairie habitats near Ft. Collins, Colorado (deCalesta 1979). In north-central Colorado, scarlet globemallow was important in desert cottontail spring and summer diets. For 2 years, the frequency of occurrence in summer diets was 67 to 78% and composition was 10 to 17%. Based on 1 year of spring data, frequency was 83% and composition was 12% (deCalesta 1979). In buffalograss-blue grama grasslands of the Texas High Plains where relative frequency of scarlet globemallow was 3.4%, the relative dry weight of scarlet globemallow in pellets was 2.6% for black-tailed jackrabbits and 2.4% for desert cottontails (Flinders and Crawford 1977).
A variety of rodents feed on scarlet globemallow vegetation and seeds. In field experiments in northwestern Nevada, Heteromyidae rodents preyed heavily on provided scarlet globemallow seed (Veech 2000). When small mammal summer diets were evaluated in chained and unchained twoseed pinyon-oneseed juniper vegetation in the Piceance Basin, Colorado, relative density of scarlet globemallow was 12% of bushy-tailed woodrats (Neotoma cinerea) and 3% of golden-mantled ground squirrel (Spermophilus lateralis) diets. At the study site, total cover of all grasses and forbs was 5% (Haufler and Nagy 1984). Average percent dry weight of scarlet globemallow plant matter in rodent stomachs was 14% for northern grasshopper mice (Onychomys leucogaster), 5% for thirteen-lined ground squirrels (Spermophilus tridecemlineatus), 3% for deer mice (Peromyscus maniculatus), and 0.4% for Ord’s kangaroo rats (Dipodomys ordii) in short-grass prairie at the Pawnee National Grassland based on plant matter recovered from the stomachs (Flake 1973). In shortgrass prairie at Colorado’s Central Plains Experimental Range, scarlet globemallow was the third most important food of northern pocket gophers (Thomomys talpoides). It was consumed most in summer and made up a high of 33% of the stomach contents from June collections. It received little to no use from December through April in the study area where it comprised 3.2% of the total vegetation cover (Vaughan 1967). Scarlet globemallow was preferred by plains pocket gophers (Geomys bursarius) in blue grama grassland in Washington County, Colorado. Although just 0.4% of the total vegetation cover in the study area, scarlet globemallow frequency in diets was 13% based on analysis of 228 stomachs over a year. Use was greatest in spring and fall (Myers and Vaughan 1964).
When diets of grazers on the Central Plains Experimental Range were compared, scarlet globemallow made up the greatest percentage of diets in fall for prairie dogs (15%), in the fall (28%) and winter (15%) for desert cottontails, and in summer (11%) for cattle (Hansen and Gold 1977).
Birds. Scaled quail (Callipepla squamata) selected scarlet globemallow in summer, winter, and fall over 3 years in the Rolling Plains of Oldham County, Texas (Ault and Stormer 1983).
Livestock. Both domestic sheep and cattle eat scarlet globemallow. Sheep use can be heavy in spring and fall, cattle use is common in the summer and may vary with grazing pressure, moisture availability, and associated vegetation.
Use of scarlet globemallow by sheep ranged from a low of 2% in a rangeland setting to a high of 81% in an experimental pasture setting (Rice et al. 1971; Rumbaugh et al. 1993b). In short-grass rangelands near Laramie, Wyoming, 2 to 5% of sheep diets were composed of scarlet globemallow during monitoring from June 7 to August 22 (Rice et al. 1971). In pastures with a mix of crested wheatgrass (Agropyron cristatum), alfalfa (Medicago sativa), and scarlet globemallow, sheep consumed 78% of scarlet globemallow in fall and 81% in spring. Fall and spring use were evaluated for 2 years and use was heavy throughout the pastures. Mortality of scarlet globemallow plants was 41% in grazed and 4% in ungrazed areas (P < 0.01) (Rumbaugh et al. 1993b).
Scarlet globemallow was seasonally important to both sheep and cattle grazing the zone between short-grass steppe and woodland vegetation at the Fort Stanton Research Station in New Mexico. Although not abundant in the study area, scarlet globemallow made up a large proportion of cattle diets in June and July and sheep diets in June, July, and October (Table 2; Thetford et al. 1971).
Table 2. Average plant composition (% dry weight) of scarlet globemallow in sheep and cattle diets at Fort Stanton in Lincoln County, NM (Thetford et al. 1971).
|
Month |
April |
June |
July |
August |
October |
January |
|
———–% dry weight———- |
||||||
|
Sheep |
3.6 |
16.0 |
7.7 |
2.8 |
16.1 |
3.5 |
|
Cattle |
0.5 |
14.2 |
29.3 |
2.2 |
3.8 |
0.2 |
Scarlet globemallow use by cattle has been reported from North Dakota through New Mexico and Texas. Most studies report heaviest use in the summer and fall. At the Dickinson Experiment Station in North Dakota, feeding on scarlet globemallow was inconsistent. At five mixed-grass prairie sites with short-duration or repeated season-long use, grazing of scarlet globemallow resulted in plant disappearance of 0 to 13% (Kirby et al. 1986). At the same site, cattle selected scarlet globemallow in proportion to its availability with short-duration grazing (5 days, 3-4 times/yr) and was preferred by cattle in fall (Pessin et al. 1986). Scarlet globemallow made up a high of 12% of summer diets at the Central Plains Experimental Range based on stomach and fecal samples collected over 2 years (Vavra et al. 1978). In short-grass prairie in Lubbock County, Texas, scarlet globemallow was prevalent in steer diets in both short-duration and continuously grazed sites (Pitts and Bryant 1987).
Use of scarlet globemallow by cattle may decline in normal moisture years, with increased grazing pressure, at sites with fourwing saltbush (Atriplex canescens), or low-nitrogen sites. At the Central Plains Experimental Range, summer use of scarlet globemallow was 15% in lightly grazed and 11% in heavily grazed areas. Significantly more scarlet globemallow was consumed in a normal than a dry summer (P < 0.1), and it was the fifth most preferred food (Vavra et al. 1977). On the same Experimental Range, cattle diets had notably more scarlet globemallow diets in shortgrass range (11–13% of summer diets) lacking saltbush than in range with fourwing saltbush, a major constituent of cattle diets (Shoop et al. 1985). At New Mexico’s Fort Stanton Research Station, scarlet globemallow was mostly consumed by cattle in June and July. In comparisons between nitrogen added and control sites there was “some indication” of increased preference and palatability with fertilization (Havstad et al. 1979).
Grazing response. Disturbances and biomass removal from the activities of black-tailed prairie dogs and livestock are typically tolerated by scarlet globemallow. In many cases, scarlet globemallow abundance was greater on grazed than undisturbed sites.
Scarlet globemallow abundance was unchanged or increased with black-tailed prairie dog grazing and disturbances. In mixed-grass prairie in northeastern Montana, biomass of scarlet globemallow was not significantly different on sites colonized by black-tailed prairie dogs and uncolonized sites (Johnson-Nistler et al. 2004). In comparisons of vegetation inside and outside black-tailed prairie dog towns at two shortgrass sites in central Colorado, scarlet globemallow cover was less than 0.5% different. Researchers concluded plants were not adversely affected by grazing and burrowing by black-tailed prairie dogs (Bonham and Lerwick 1976). In the Cimarron National Grassland, Kansas, frequency of scarlet globemallow was greater on colonized than uncolonized short- and mid-grass plots, and differences were significant between colonized (35%) and uncolonized (14%) short-grass plots (P < 0.05; Winter et al. 2002). In semi-arid grasslands near Denver, Colorado, scarlet globemallow occurred as a dominant on more than 40% of colonized sites but on less than 5% of uncolonized areas (Magle and Crooks 2008).
In Badlands National Park, South Dakota, scarlet globemallow was lower at sites used by both black-tailed prairie dogs and bison than at sites without bison or without either species. Vegetation comparisons were made in 3-year-old exclosures in northern mixed-grass prairie. Cover of scarlet globemallow was 6.75% in completely protected, 7.25% in black-tailed prairie dog only, and 3.25% in unprotected (bison and black-tailed prairie dog accessible) (Fahnestock and Detling 2002). In captive feeding experiments, only 7% of scarlet globemallow seeds fed to bison were recovered undamaged. None of these undamaged seeds germinated suggesting that passage through the digestive tract was insufficient to break dormancy (Gokbulak 2002).
Studies suggest scarlet globemallow does persist and often increases with grazing, but plant response can vary with grazing intensity, moisture conditions, and soils. Although referred to as an increaser with grazing (Tomanek and Albertson 1957), when sheep-grazed and mostly protected big sagebrush grasslands were compared in Sweetwater County, Wyoming, scarlet globemallow occurred on protected but not grazed sites (Marquiss and Lang 1959). In blue grama-buffalograss communities at the Pawnee National Grassland, scarlet globemallow abundance was greater on lightly (1 animal unit/12.1 ha/mo) than heavily (3 animal units/12.1 ha/mo) cattle-grazed sites (Schwartz and Nagy 1976). Canopy cover of scarlet globemallow was significantly greater in grazed (0.27%) than ungrazed little bluestem steppe (0.05%) on the Pawnee National Grassland in eastern Colorado (P < 0.05). Areas lightly grazed by livestock resulting in 30 to 35% removal of annual standing biomass (Milchunas and Noy-Meir 2004). On the Central Plains Experimental Range, density of scarlet globemallow decreased on grazed and increased on ungrazed sites in a dry year. Density differences on grazed and ungrazed sites were less pronounced in a wet year. Grazed sites were subjected to heavy, long-term cattle grazing (Milchunas et al. 1989). In salt desert shrublands in western Colorado, frequency differences on grazed and ungrazed sites were more pronounced on shale than on sandstone or mixed shale-sandstone soils. The study area was heavily grazed by cattle and sheep during winter and spring. Exclosures protected areas from livestock for 10 years. On shale soils, frequency of scarlet globemallow was 10% on grazed and 2% on protected plots. On sandstone soils, frequency was 54% on grazed and 52% on protected. On mixed soils, frequency was 26% on grazed plots and 12% on protected plots (Turner 1971).
Nutritional Value
In controlled experiments, including forbs (50:50 mix of scarlet globemallow and leatherweed [Croton pottsii]) with low-quality forage diets was, in most instances, comparable to including alfalfa. Results suggested that maintaining palatable forbs on rangeland sites should reduce the need to supply cattle with protein when grasses are dormant (Arthun et al. 1992b). Additional analyses revealed few changes in ruminal digesta kinetics and fermentation patterns when cattle diets were supplemented with forbs rather than alfalfa (Arthun et al. 1992a).
In a blue grama-buffalograss community in northeastern Colorado, nitrogen content of scarlet globemallow was greatest from May (2.8%) through June (2.4%) and decreased in the following months through October (1.5%). These measurements were made on samples collected from May through October 1995 through 1999 (Augustine et al. 2013).
Detailed nutritional analyses are available from spring and fall samples of scarlet globemallow growing in southern Idaho (Table 3; Rumbaugh et al. 1993a) and in Hays, Kansas (Table 4; Runyon 1947).
Table 3. Nutritional content (evaluated and averaged for fall and spring) of scarlet globemallow plants growing near Kimberley, Idaho. Pastures with scarlet globemallow growing with alfalfa or crested wheatgrass meet the dietary requirements of sheep and cattle (Rumbaugh et al. 1993a).
|
Measurement |
Microminerals (µg/g) |
Macrominerals mg/g |
||||||||
|
Element |
Cu |
Fe |
Mn |
Na |
Zn |
Ca |
Mg |
N |
P |
K |
|
Leaves |
18 |
1230 |
61 |
1050 |
30 |
24 |
5.2 |
32 |
3.2 |
18 |
|
Stems |
21 |
460 |
31 |
810 |
44 |
16 |
4.7 |
23 |
2.4 |
18 |
Table 4. Chemical composition of scarlet globemallow growing in Hays, Kansas. Plant samples are averages for summer and fall collections in 1 year (Runyon 1947).
|
Chemical composition (%) |
Ash |
Ca |
P |
Fat |
Fiber |
Crude protein |
N-free extract |
|
Scarlet globemallow |
11.6 |
3.5 |
0.4 |
14.9 |
12.2 |
7.8 |
49.6 |
A sample of scarlet globemallow seeds contained 7.4% oil and 12.9% protein (Earle and Jones 1962 cited in RGB Kew 2018). Seeds collected near Albuquerque, New Mexico, had 19.2 J/mg of energy and were 3.8% nitrogen (Henderson 1990).
Ethnobotany
Scarlet globemallow was used medicinally and ceremonially by Indians. Blackfoot Indians chewed the plant into a paste, which was used as a healing salve for sores and wounds. Blackfoot medicine men would apply the paste to hands and arms, which would allow the medicine men to retrieve items from boiling water unharmed (Johnston 1970). The Lakota, Dakota, and Navajo used scarlet globemallow to treat pain. Plants (often roots) were chewed into a poultice to cool and heal sores and wounds. The Comanche used an infusion of the plant to ease swelling (Moerman 2003). Acoma and Laguna Indians used scarlet globemallow roots as a medicine for pregnant women (Swank 1932). The Kayenta Navajo dried the plant into a powder to dust sores. The plant was also used as a lotion for skin ailments, a tonic to improve appetite, and a treatment for hydrophobia (Wyman and Harris 1951). The Navajo referred to scarlet globemallow as a medicine plant with gummy roots. An infusion of the plant was used as a remedy for diseases produced by witchcraft. Scarlet globemallow roots were chewed in times of famine and used to stop bleeding (Elmore 1944).
The northern Cheyenne of Montana and Kayenta Navajo used scarlet globemallow in ceremonies (Wyman and Harris 1951; Hart 1981). Plants were used as a ceremonial fumigant ingredient, said to give strength to singers, and referred to as a ‘life medicine’ by the Navajo (Wyman and Harris 1951).
The Gosiute of Utah pounded scarlet globemallow into a gummy paste and applied it over the insides of earthen dishes to fill small holes. Once the paste hardened, the dish was left with a smooth surface (Chamberlin 1911).
Horticulture
Scarlet globemallow is listed as a water wise native plant for use in Intermountain landscapes (Mee et al. 2003). It is recommended for USDA Plant Hardiness Zones 6 to 8, where annual precipitation averages 6 to 10 or more in (150–250 mm) (St. John and Ogle 2009).
Revegetation Use
Scarlet globemallow has many traits that make it a good choice in revegetation and restoration of communities throughout the Great Plains and Intermountain West, where annual precipitation averages 7 in (180 mm) or more (USDA NRCS IDPMC 2005; Ogle et al. 2011). The species tolerates disturbed and drought conditions and when a seed source is present, often colonizes and stabilizes soils on open sites (NRCS IDPMC 2005). The species is also important to a diversity of wildlife species, insects, and pollinators (Ley et al. 2007; Ogle et al. 2011). Scarlet globmallow was rated as good to excellent for ease of handling, ease of seeding, ease of transplanting, seedling growth rate, final establishment, persistence, grazing tolerance, compatibility with other species, and soil stability in big sagebrush, shadscale saltbush (Atriplex confertifolia), black greasewood (Sarcobatus vermiculatus), blackbrush, mountain brush, and pinyon-juniper (Stevens and Monsen 2004). Scarlet globemallow was reported on abandoned untreated and reclaimed mine sites, roadcuts, and similar sites throughout its range (Sieg et al. 1983; Fisher et al. 1990; Weaver et al. 1993).
On abandoned mine sites, scarlet globemallow is often an early colonizing species irrespective of reclamation treatments. In Carter County, Montana, scarlet globemallow occurred on unreclaimed (12–28 yrs old) and reclaimed (5–12 yrs old) bentonite mine spoils, even though it was absent from the reclamation seed mix (Sieg et al. 1983). Natural colonization also occurred on a recontoured surface mine near Kemmerer, Wyoming, where topsoil treatments were implemented but scarlet globemallow was not planted or seeded (Hatton and West 1987). Scarlet globemallow also colonized coal mine spoils about 10 years after abandonment in northwestern New Mexico (Wagner et al. 1978). On the Cibola National Forest in New Mexico, scarlet globemallow naturally colonized abandoned uranium mine spoils during reclamation, although not intentionally planted or seeded. It occupied 2% of the experimental area 4 years after initiation of reclamation, which included fertilization, addition of wood chips, and planting oneseed juniper (Fisher et al. 1990). Scarlet globemallow recovered or colonized soon after the severe disturbance caused by a salt water blow out associated with oil drilling operations in western North Dakota. The blow out occurred in April 1982. Density of scarlet globemallow in the damaged area was 0.1 plant/ft² (1.5 plants/m²) in July 1982 and 0.2 plant/ft² (2.6/m²) in July 1984. Plants were absent from reclaimed sites treated with calcium chloride (Halvorson and Lang 1989).
Because of its natural occurrence along roadsides and general wide adaptability or tolerance, scarlet globemallow was listed as a useful roadside revegetation species in grasslands and forest types in Montana (Meier and Weaver 1997). When roadside cuts and adjacent undisturbed, near climax vegetation were compared in the northern Rocky Mountains, scarlet globemallow occurred on both roadsides and in climax grasslands (Weaver et al. 1993).
Scarlet globemallow invaded experimental revegetation plots in the Piceance Basin, Colorado (Grygiel et al. 1984). Sites were scalped, ripped 12 in (30 cm) deep, and seeded with mixes of native and nonnative grasses, forbs, and shrubs in 1976. By 1979, scarlet globemallow seedlings and relatively small, mature plants occurred as relatively large clumps or islands with multiple plants. Over the next 2 years, the islands were fragmented into more but smaller clumps. Plants were described as “prolifically rhizomatous”. Young and parent plants were connected by rhizomes up to 3.3 ft (1 m) long. Researchers considered scarlet globemallow an aggressive invader of open sites but not an aggressive competitor with established plants. Though abundance of scarlet globemallow decreased as competition with maturing neighbor plants increased, it remained an important vegetation component (Grygiel et al. 1984).
Developing A Seed Supply
For restoration to be successful, the right seed needs to be planted in the right place at the right time. Coordinated planning and cooperation is required among partners to first select appropriate species and seed sources and then properly collect, grow, certify, clean, store, and distribute seed for restoration (PCA 2015).
Developing a seed supply begins with seed collection from native stands. Collection sites are determined by current or projected revegetation requirements and goals. Production of nursery stock requires less seed than large-scale seeding operations, which may require establishment of agricultural seed production fields. Regardless of the size and complexity of any revegetation effort, seed certification is essential for tracking seed origin from collection through use (UCIA 2015).
Seed Sourcing
Because empirical seed zones are not currently available for scarlet globemallow, generalized provisional seed zones developed by Bower et al. (2014), may be used to select and deploy seed sources. These provisional seed zones identify areas of climatic similarity with comparable winter minimum temperature and aridity (annual heat:moisture index). In Figure 5, Omernik Level III Ecoregions (Omernik 1987) overlay the provisional seeds zones to identify climatically similar but ecologically different areas. For site-specific disturbance regimes and restoration objectives, seed collection locations within a seed zone and ecoregion may be further limited by elevation, soil type, or other factors.
In a study to determine the correlations between temperature and grassland vegetation change as a way to inform potential changes associated with future climate, researchers found scarlet globemallow was negatively associated with minimum winter temperatures (P = 0.038). Using long-term data (23 years) collected from shortgrass steppe at the Central Plains Experimental Range, Colorado, researchers found that as winter minimum air temperatures increased, production of scarlet globemallow decreased (Alward et al. 1999).
The Western Wildland Environmental Threat Assessment Center’s (USFS WWETAC 2017) Threat and Resource Mapping (TRM) Seed Zone application provides links to interactive mapping features useful for seed collection and deployment planning. The Seedlot Selection Tool (Howe et al. 2017) can also guide restoration planning, seed collection, and seed deployment, particularly when addressing climate change considerations.
Occurrence Map
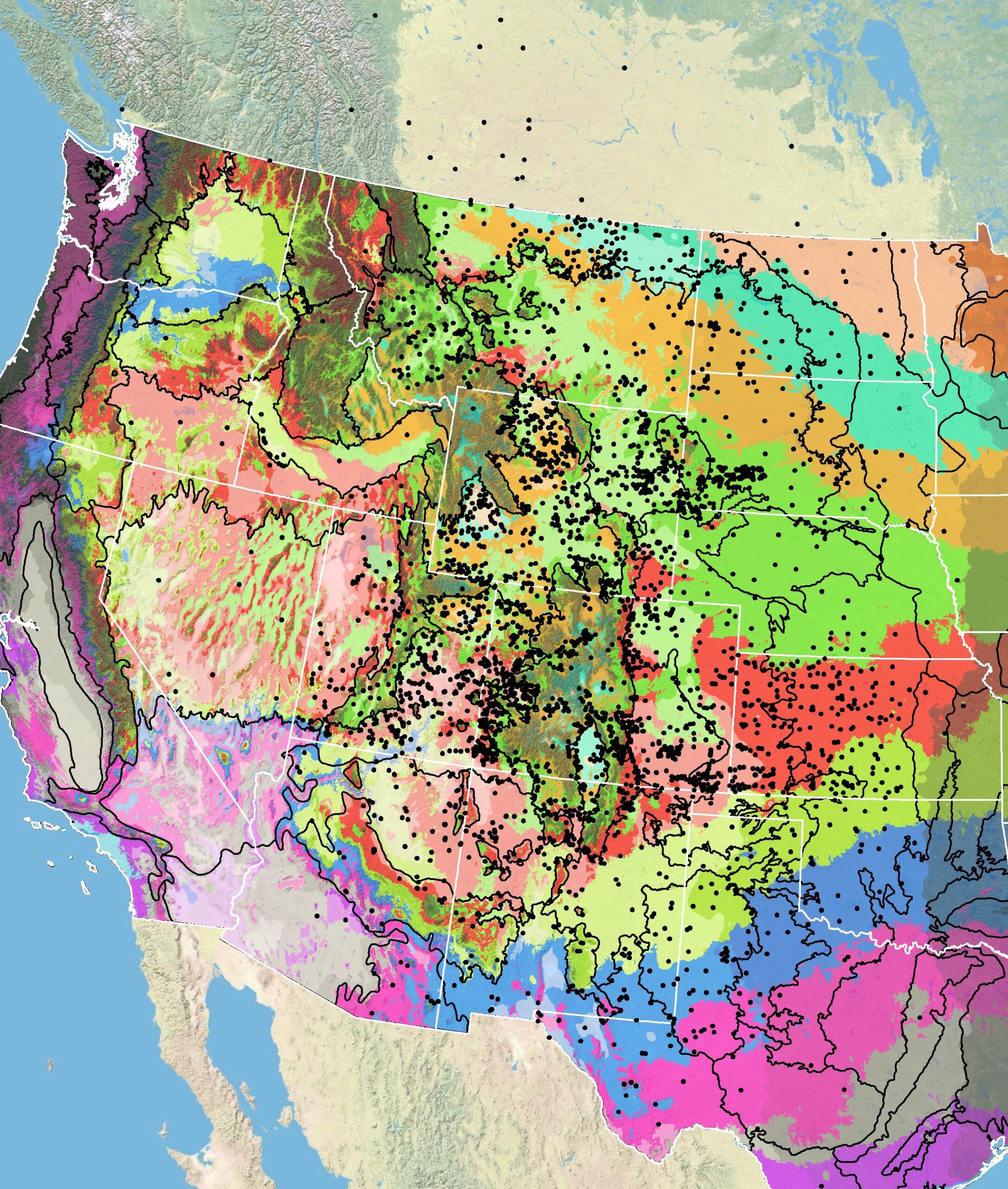
Figure 5. Distribution of scarlet globemallow (black circles) based on geo-referenced herbarium specimens and observational data from 1849-2016 (CPNWH 2017; SEINet 2017; USDI USGS 2017). Generalized provisional seed zones (colored regions) (Bower et al. 2014) are overlain by Omernik Level III Ecoregions (black outlines) (Omernik 1987; USDI EPA 2018). Interactive maps, legends, and a mobile app are available (USFS WWETAC 2017; research.fs.usda.gov/pnw/products/dataandtools/seed-zone-webmap). Map prepared by M. Fisk, USDI USGS
Releases
There is one germplasm release, Scarlet Globemallow ARS 2936 (Stevens et al. 1996). It came from seed collected in northern Idaho and was selected for its spread and stem production from rhizomes (Pendery and Rumbaugh 1993). The ARS-2936 accession was grown in a forage pasture study in Kimberly, Idaho (Rumbaugh et al. 1993c). This accession produced twice as much forage as other accessions, and its rhizome growth lead to full colonization of some of the 0.4 ft² (4 m²) plots within 6 months of transplanting even when growing with crested wheatgrass. In non-competitive growth plots near Logan, Utah, rhizomes of 1-year-old ARS 2936 plants extended a minimum of 18 in (45 cm) and a maximum of 44 in (113 cm). The release was suggested as useful for forage, soil stabilization, and highway revegetation or beautification in semi-arid to arid sites in the Great Plains and Rocky Mountains (Rumbaugh et al. 1993c).
Wildland Seed Collection
Scarlet globemallow is one of the earliest flowering globemallows (La Duke 2016). Indeterminate flowering and seed production mean collections may be made at a single site for weeks (Hewitt 1980) and across sites for months (USDI BLM SOS 2017). Seed is released once schizocarps are fully mature, so careful monitoring once plants begin to flower is needed to maximize wildland seed collections (Fig. 6) (Shock et al. 2015). In 1997, between 50 and 1,000 lbs (20–450 kg) pure live seed (PLS) of wild-collected scarlet globemallow seed was available (McArthur and Young 1999).

Figure 6. Scarlet globemallow with ripe schizocarps. Photo: BLM UT030 SOS.
Wildland Seed Certification
Verification of species and tracking of geographic source is necessary whether wildland seed is collected for immediate project use or as stock seed for cultivated increase. This official Source Identification process can be accomplished by following procedures established by the Association of Official Seed Certifying Agencies (AOSCA) Pre-Variety Germplasm Program (UCIA 2015; Young et al. 2020). Wildland seed collectors should become acquainted with state certification agency procedures, regulations, and deadlines in the states where they collect.
If wildland-collected seed is to be sold for direct use in ecological restoration projects, collectors must apply for Source-Identified certification prior to making collections. Pre-collection applications, site inspections, and species and seed amount verification are handled by the AOSCA member state agency where seed collections will be made (see listings at AOSCA.org).
If wildland seed collected by a grower or private collector is to be used as stock seed for planting cultivated seed fields or for nursery propagation (See Agricultural Seed Field Certification section), detailed information regarding collection site and collecting procedures must be provided when applying for certification. Photos and herbarium specimens may be required. Germplasm accessions acquired within established protocols of recognized public agencies, however, are normally eligible to enter the certification process as stock seed without routine certification agency site inspections. For contract grow-outs, however, this collection site information must be provided to the grower to enable certification.
Collection Timing
Flowering of scarlet globemallow can occur from March through October throughout its broad range (Kemp 1983; Holmgren et al. 2005). Timing of flowering and seed production can vary with site conditions, elevation, and climate (Wasser 1982; Kemp 1983), but spring and summer flowering is most common for both varieties (Craighead et al. 1963; Wasser 1982; White 1995; La Duke 2016; USDI BLM SOS 2017). Additional flowering dates and information are available in the Phenology section.
Of 37 total seed harvests made by BLM SOS crews, 22 (or about 60%) were made in July (Fig. 7). Collections spanned 12 years and elevations of 4,000 to 7,700 ft (1,200–2,300 m) in Colorado, Wyoming, and Utah. The earliest collection was made on June 5, 2016 from 4,867 ft (1,483 m) in Mesa County, Colorado. The latest collection date was August 18, 2009 from 4,722 ft (1,439 m) in Big Horn County, Wyoming (USDI BLM SOS 2017). Seed maturity is also common in August throughout the range of the species (Belcher 1985). For scarlet globemallow plants growing in exclosures in sagebrush-short grass vegetation in northern Colorado, fruit development and seed shatter occurred in August during 2 years of observations (Menke and Trlica 1981).

Figure 7. Scarlet globemallow with open schizocarps shedding or having already shed seed. Photo: BLM UT030 SOS.
In the northern Chihuahuan Desert, scarlet globemallow produced flowers and seed twice a year between July 1978 and June 1980 (Kemp 1983). Flowering occurred in August 1978, and fruits were produced in September. In 1979, the first flowering began in early April, and first fruits were produced in March. A second flowering occurred in August, and fruits were produced again in September. In 1980, flowering occurred in mid- to late April, and fruits were produced in mid- to late March. Precipitation in the study area was most abundant and reliable in late summer (July-September), and 30 or more days without precipitation were common in the spring (March-May) and fall (October-December). During this study, precipitation was abundant in late summer and fall 1978, and there was an extended drought from September to December 1979 (Kemp 1983).
Collection Methods
Wildland seed can be harvested by hand by stripping inflorescences or knocking fruits and seeds into a container (Wasser 1982; Jorgensen and Stevens 2004). Protection of the skin, eyes, and airways is advisable because of the irritating stellate hairs covering scarlet globemallow herbage, fruits, and seeds (Fig. 8) (St. John and Ogle 2009). In dense stands occurring in suitable terrain, seed can be mechanically harvested with a reel-type harvester (Jorgensen and Stevens 2004).
Scarlet globemallow seeds are subject to insect damage (LBJWC 2018) and careful inspection of populations for seed damage can help maximize harvesting efforts.
Several collection guidelines and methods should be followed to maximize the genetic diversity of wildland collections: 1) collect seed from a minimum of 50 randomly selected plants; 2) collect from widely separated individuals throughout a population without favoring the most robust or avoiding small stature plants; and 3) collect from all microsites including habitat edges (Basey et al. 2015). General collecting recommendations and guidelines are provided in online manuals (e.g., ENSCONET 2009; USDI BLM SOS 2021).
It is critical that wildland seed collection does not impact the sustainability of native plant populations. Collectors should take no more than 20% of the viable seed available at the time of harvest (USDI BLM SOS 2021). Additionally, care must be taken to avoid the inadvertent collection of weedy species, particularly those that produce seeds similar in shape and size to those of scarlet globemallow.
Post-Collection Management
Thorough drying of harvests and protection from insect damage are necessary for scarlet globemallow.


Figure 8. Collection of scarlet globemallow seeds and schizocarps before cleaning (left). Photo: BLM WY050 SOS. Single mericarp and seed (right). Photo: BLM WY932A SOS.
Seed Cleaning
Dry seed can be cleaned by processing it through a debearder, air screen separator, and gravity table (Stevens et al. 1996; Jorgensen and Stevens 2004).
The USFS Bend Seed Extractory used the following procedure to clean a small lot (2.3 lbs [1 kg]) of wildland seed collected from Montrose County, Colorado:
- Process using a laboratory brush machine with a #18 mantel and medium speed.
- Air-screen using an office Clipper with a size 8, round, top screen and size 1/20, round bottom screen, medium speed, and medium air.
This cleaning procedure yielded 315,000 seeds/lb (694,000 seeds/kg). Seed fill was 90%, and purity was 95% (Barner 2009). Seed purity can be increased to 90% through use of a gravity table in the cleaning process (Wasser 1982; Stevens et al. 1996).
Seed Storage
Scarlet globemallow seed can be stored for 16 years or more (Stevens et al. 1996; Jorgensen and Stevens 2004). Seed is orthodox and there was no loss of viability when dry seed was stored at -4 °F (-20 °C) for 1.7 years (RGB Kew 2018). At the Bend Seed Extractory, scarlet globemallow seed was stored at 33 to 38 °F (0.5–3 °C) (Barner 2009).
Seed Testing
There is no Association of Seed Analysts (AOSA) germination protocol for scarlet globemallow (AOSA 2016), though AOSA guidelines for tetrazolium chloride (TZ) viability testing of the Sphaeralcea genus are available. Seed is prepared by an overnight soak at 68 to 77 °F (20–25 °C) followed by staining with 0.1 or 1% TZ concentrations. Seed is not viable if any part of the embryo or endosperm is unstained. Clipping of the seed coat is recommended for hard seeds (AOSA 2010).
Quick estimates of seed fill can be made using the ‘pop test’ described by Tilley et al. (2011). This test uses a hot plate to heat seeds until the moisture contained within the seed is converted to gas, breaks the seed coat, and produces a pop. Germination and viability tests were not done on seeds that popped.
Purity Testing
Routine seed purity analysis requires 2 g of scarlet globemallow seed. For noxious weed counts, 20 g of seed is required (Belcher 1985).
Germination Biology
Scarification is required for scarlet globemallow germination, although seed origin and seed storage conditions may affect germination requirements. Stevens and Jorgensen (2004) suggest afterrippening seed for a month and then scarifiing or cold stratifing for 1 to 3 months for good germination. While many studies show that scarification is required for high levels of germination of Sphaeralcea spp., boiling water treatments were considered the only operationally viable treatments for large seed lots (Kildisheva et al. 2013).
Complete germination occurred when seed was scarified by chipping or partially removing the seed coat, exposed to a 8/16-hour light/dark cycle, and exposed to fluctuating temperatures of 77/50 °F (25/10 °C) (RBG Kew 2018).
For wildland seed collected from Uinta County, Utah, germination levels were high (≥70%) for scarified seed treated to early spring conditions (4 wks at 52/34 °F [11/1 °C]) and scarified or unscarified seed treated to winter (12 wks at 34 °F [1 °C]) followed by early spring conditions (Kramer and Foxx 2016). Seeds were incubated in growth chambers with a 12-hr light/dark cycle. Germination was lower (< 50%) when seeds were exposed to warmer temperatures (59/41 °F [15/5 °C] and 68/50 °F [20/10 °C]) regardless of scarification and even when low temperatures exposure (winter or early spring) preceded warmer temperature exposure (Kramer and Foxx 2016).
Scarification can improve germination of scarlet globemallow seed. A 20-minute soak in sulfuric acid produced germination levels of about 25%, which was significantly greater than that of seeds soaked in water (~7%) (P < 0.05; Meyer and Vernon 2008). Acid-scarified seed (77%) germinated significantly better than untreated seed (5%) (P < 0.05), but mechanical scarification in a metal drum lined with sand paper for 10 seconds or more resulted in embryo damage and seedling death within 7 days (Roth et al. 1987). Mechanical scarification did significantly improve germination of scarlet globemallow seed obtained from a grower in Wisconsin (Dunn 2011). Seed was scarified by rubbing it down a 6-in (15 cm) length of 320-grit aluminum oxide emery cloth. A proportion of scarified and control seed was then stratified in moist media at 39 °F (4 °C) for 30 days before treated and untreated seeds were put in a greenhouse (68°F [20 °C]). Germination was 4.7% for stratified only seed. Germination was significantly greater for scarified only seed (51.3%) and scarified and stratified seed (85.3%) (P < 0.05). Researchers concluded the main factor that prevented germination was the impermeable seed coat but that embryos appeared partially or conditionally dormant (Dunn 2011).
Wildland Seed Yield And Quality
Post-cleaning seed yield and quality of scarlet globemallow seed lots collected in the Intermountain region are provided in Table 5 (USFS BSE 2017). The results indicate that scarlet globemallow seed can generally be cleaned to high levels of purity and fill and that viability of fresh seed is variable. Seed weights reported in Table 5 were similar to those reported (125,976–215,959 seeds/lb) by several others (Stevens 1957; USFS GBNPP 2014; RGB Kew 2018), but much lower than those reported by Stevens et al. (1996) (500,000 seeds/lb [1,102,000 seeds/kg]) and Belcher (1985) (681,000 seeds/lb [1,362,000 seeds/kg]).
Table 5. Seed yield and quality of scarlet globemallow seed lots collected in the Intermountain region, cleaned by the Bend Seed Extractory, and tested by the Oregon State Seed Laboratory or the USFS National Seed Laboratory (USFS BSE 2017).
|
Seed lot characteristic |
Mean |
Range |
Samples (no.) |
|
Bulk weight (lbs) |
2.07 |
0.11–24.5 |
21 |
|
Clean weight (lbs) |
0.75 |
0.02–9.95 |
21 |
|
Clean-out ratio |
0.36 |
0.05–1.23 |
21 |
|
Purity (%) |
98 |
97–99 |
21 |
|
Fill (%)¹ |
90 |
70–99 |
21 |
|
Viability (%)² |
89 |
71–98 |
20 |
|
Seeds/lb |
162,099 |
128,100–226,003 |
21 |
|
Pure live seeds/lb |
142,532 |
109,483–183,831 |
20 |
¹100 seed X-ray test
²Tetrazolium chloride test
Marketing Standards
Seed purity, viability, and germination specifications vary with revegetation plans. Purity needs are highest for precision seeding equipment used in nurseries, while some rangeland seeding equipment handles less clean seed quite well.
Acceptable seed viability for marketed scarlet globemallow seed is 90% and seed purity is 80% (Jorgensen and Stevens 2004). Acceptable germination of marketed seed was reported as 20 to 80% (Wasser 1982; Jorgensen and Stevens 2004). In 1997, a pound of scarlet globemallow PLS was $25 to $35 and about 1,000 to 1,500 lbs (2,200–3,300 kg) PLS of wildland-collected seed was available (McArthur and Young 1999).
Agricultural Seed Production
Scarlet globemallow seed production plots were grown successfully at a site north of Vernon, Utah, (Jones and Whittaker 2011) and at Oregon State University’s Malheur Experiment Station (OSU MES) (Shock et al. 2015).
Agricultural Seed Certification
In order to minimize genetic changes in specific accessions of native species when increased in cultivated fields, it is essential to track the geographic source and prevent inadvertent hybridization or selection pressure. This is accomplished by following third party seed certification protocols for Pre-Variety Germplasm (PVG) as established by the Association of Official Seed Certification Agencies (AOSCA). AOSCA members in the U.S., Canada, and other countries administer PVG requirements and standards that track the source and generation of planting stock. Field and cleaning facility inspections then monitor stand establishment, proper isolation distances, control of prohibited weeds, seed harvesting, cleaning, sampling, testing, and labeling for commercial sales (UCIA 2015; Young et al. 2020).
Seed growers apply for certification of their production fields prior to planting and plant only certified stock seed of an allowed generation (usually less than four). The systematic and sequential tracking through the certification process requires preplanning, knowing state regulations and deadlines, and is most smoothly navigated by working closely with state certification agency personnel. See the Wildland Seed Certification section for more information on stock seed sourcing.
Site Preparation
Seed production fields at OSU MES were established in weed free plots (Shock et al. 2015).
Seed Pretreatments
Boiling water was poured on scarlet globemallow seed prior to direct seeding plots in Utah. This treatment was successful in melting away the water-impermeable seed coat, allowing the seeds to take up water and germinate. Dense stands were produced using this seed pretreatment procedure (Jones and Whittaker 2011).
Weed Management
Mechanical or chemical treatment of weeds prior to seeding and hand weeding as needed after planting is recommended for seed production fields (Stevens et al. 1996). At OSU MES, hand weeding was used to control competition after planting in seed production plots (Shock et al. 2015).
Seeding
General recommendations for scarlet globemallow production plots include a seeding rate of 20 to 50 PLS/linear ft of row, row spacing of 28 to 36 in (71–91 cm), and seeding depths of 0.13 to 0.25 in (3–6 mm) (Pendery and Rumbaugh 1993; Stevens et al. 1996). Fall or winter seeding or use of scarified seed at sites with sandy loam to clay-textured soils is also recommended (Pendery and Rumbaugh 1993; Stevens et al. 1996). In seed productions plots at OSU-MES, 30 seeds were planted per foot of row (Shock et al. 2015).
Effects of seeding depth and soil type on emergence and seedling survival were evaluated in a greenhouse study. Seedling survival to 45 days was significantly greater in sandy loam (71%) than clay loam (32%) field-collected soils regardless of seeding depth (P = 0.009). Scarlet globemallow seed came from a 5,029-ft (1,533 m) elevation site in Carbon County, Utah, and was seeded at depths 0 to 1-in (2.5 cm). Emergence was greatest for seeds planted at 0.25-in (6 mm) depths but not significantly greater than emergence from 0, 0.3 cm, 0.12 cm, or 1-in (2.5 cm) depths in sandy loam soils but was significantly greater (P ≤ 0.10) than emergence from seeds planted 1 in (2.5 cm) deep in clay loam soils. Seedling survival through day 45 was greatest for seeds planted 0.25 in (6 mm) deep in both soil types (Rawlins et al. 2009).
Fall seeding is recommended for scarlet globemallow. When seed was planted in April, stand emergence was poor. Spring sowing involved use of cold-stratified seed that was soaked in distilled water, rinsed with a bleach solution, and cold-stored for about 2.5 months at 34 °F (1 °C). When seed was fall planted, stand emergence was good (Shock et al. 2012, 2015).
Establishment And Growth
For seed production, low to moderate fertilization of scarlet globemallow has been recommended (Stevens et al. 1996). In wildland conditions, however, scarlet globemallow abundance often decreased with fertilization (Goetz 1969; Samuel and Hart 1998; McLendon and Redente 1991).
When a nitrogen availability gradient was established and evaluated in disturbed and seeded plots at northwestern Colorado’s Piceance Basin, relative cover of scarlet globemallow averaged over 3 years was 0.7% in high-nitrogen, 1% in low-nitrogen and 1.5% in no-nitrogen plots (McLendon and Redente 1992). The study area was scalped to 2 in (5 cm), all vegetation was removed, and the next 14 in (35 cm) of soil was mixed to simulate disturbances created by energy development. The species seeded and seeding densities were not reported, and nitrogen was applied (89 lbs N/ac/yr [100 kg/ha/yr]) for 3 post-disturbance years (McLendon and Redente 1992).
In wildland stands, the abundance of scarlet globemallow plants was lower or unchanged with fertilization. In grasslands at the Dickinson Experiment Station, density and cover of scarlet globemallow growing in fine sand and silt loam was lower at nitrogen-fertilized (33–100 lbs/ac [37–112 kg/ha]) than unfertilized (Goetz 1969). At a shallow silt loam soil site, scarlet globemallow density was greater and cover lower at fertilized than unfertilized plots. Differences between density and cover on fertilized and unfertilized plots were not significant at any site (Goetz 1969). At the High Plains Grasslands Research Station near Cheyenne, Wyoming, scarlet globemallow frequency was nearly always greater on unfertilized than fertilized plots in mixed-grass prairie. Frequency was evaluated from 1976 to 1988, and nitrogen fertilization (20–30 lbs/ac [22–34 kg/ha]) was applied for 8 years (1974–1982) (Samuel and Hart 1998). At the Piceance Basin, biomass of scarlet globemallow was greater on control than nitrogen-treated plots in semi-arid big sagebrush (McLendon and Redente 1991). Three years following nitrogen additions in mixed grass near Cheyenne, Wyoming, frequency of scarlet globemallow was unaffected by one-time fertilizer applications of 200 to 600 lbs/ac (224–672 kg/ha) (Houston and Hyder 1975).
Irrigation
Targeted irrigation during the establishment phase and again in the spring and during the seed set or seed ripening phase have been recommended for seed production plots at sites where annual precipitation is 16 in (406 mm) or less (Stevens et al. 1996; St. John and Ogle 2009). In experimental irrigation trials at OSU MES, seed yield was increased in only a few years and not significantly with supplemental irrigation beyond that provided during the establishment phase (Table 6; Shock et al. 2015). Irrigation research findings suggested that natural rainfall in Ontario, Oregon, allowed for maximum seed production in weed-free conditions (Table 7).
Table 6. Seed yield (lbs/ac) for fall-seeded (November 2006) scarlet globemallow in response to no additional irrigation and supplemental irrigation (4 and 8 in) at Oregon State University’s Malheur Experiment Station in Ontario, OR (Shock et al. 2015).
| Year | Supplemental irrigation (inches/season*) | ||
| 0 | 4 | 8 | |
| Seed yield (lbs/ac) | |||
| 2007** | 279.8 | 262.1 | 310.3 |
| 2008 | 298.7 | 304.1 | 205.2 |
| 2009 | 332.2 | 172.1 | 263.3 |
| 2010 | 385.7 | 282.6 | 372.5 |
| 2011 | 89.6 | 199.6 | 60.5 |
* Irrigation season was from bud to seed set and water was delivered through a sub-surface drip irrigation system.
** 2007 data represents the total of three separate hand harvests. The following year’s data represent a single combine harvest.
Table 7. Natural precipitation (amounts and timing) and growing degree days at Oregon State University’s Malheur Experiment Station in Ontario, OR (Shock et al. 2015).
|
Year |
Jan-June (in) |
April-June (in) |
Growing degree days (50-86 ºF) Jan-June |
|
2006 |
9.0 |
3.1 |
1120 |
|
2007 |
3.1 |
1.9 |
1208 |
|
2008 |
2.9 |
1.2 |
936 |
|
2009 |
5.8 |
3.9 |
1028 |
|
2010 |
8.3 |
4.3 |
779 |
|
2011 |
8.3 |
3.9 |
671 |
|
Average |
5.8 |
2.7 |
1042 |
Pollinator Management
Scarlet globemallow flowers are strongly outcrossing, and bees, especially Diadasia species, are important pollinators (Pendery and Rumbaugh 1993). Specialist ground nesting Diadasia bees colonized and nested within rows of scarlet globemallow growing at the OSU MES, despite its agricultural setting (Cane 2009). Any management to encourage native bee populations in seed production areas may benefit yields (Cane 2008).
Pest Management
Scarlet globemallow is host to several fungal and insect pests, some of which have been problematic in seed production plots. Powdery mildew (Leveillula taurica) and rust (Puccinia sherardiana) were found on scarlet globemallow plants in Idaho and Oregon (Sampangi et al. 2012). Other rusts (P. recondita, P. schedonnardi) use scarlet globemallow as a host and are widely distributed throughout its range (Farr and Rossman 2017). In field studies conducted in the southwestern US, scentless plant bug nymphs (Arhyssus confuses, A. parvicornus) were considered globemallow specialists and collected from scarlet globemallow plants in Arizona and New Mexico (Wheeler 2011). Wood-boring beetles (Acmaeodera quadrivittata) were also collected from scarlet globemallow plants (Nelson and Westcott 1995).
Many insect and fungal pests have been reported in scarlet globemallow seed production plots. At OSU MES, P. sherardiana caused substantial leaf loss and reduced vegetative growth in stands in 2 of 5 years (Shock et al. 2015). Both grasshoppers and rusts were problematic at seed production fields in Utah. First-year stands were devastated by a fungal rust and failed to re-emerge the following growing season. Established stands that were heavily preyed upon by grasshoppers came back only sporadically the following growing season (Jones and Whittaker 2011).
Tarnished plant bugs (Lygus lineolaris), western plant bugs (L. hesperus), and weevils (Acanthoscelides, Apion, Anthonomus spp.) were collected from scarlet globemallow seed production fields in Colorado and Oregon. In seed fields growing near Hotchkiss, Colorado, adult seed-feeding weevils (Apionidae) were collected in suction samples collected on May 11 when the field was in full bloom. In a plant sample collected on May 21, eggs, larvae, pupae, and adults were present. Support of multiple weevil generations by scarlet globemallow’s indeterminate flowering suggest high seed damage potential. Two weevil species, Anthonomus sphaeralciae (dominant) and Apion spp., were found in 25% of flower buds examined. Management of insect predators starts with monitoring for their presence by sweep or suction monitoring for adults and close visual inspections of flowers and fruits for larvae or pupae. Treatment options are limited because there is no insecticide labeled for use, and insecticides are not a good option while plants are in flower and being visited by pollinators (Hammon and Franklin 2012; CSU Ext. 2016).
Seed Harvesting
When seeded in the fall, scarlet globemallow seed can be harvested within a year (Shock and Feibert 2016). Seed is ready for harvest when lower capsules begin to dry, which often occurs July 5 to August 5 in Utah. Harvests can be made by hand stripping or by combining (Stevens et al. 1996; St. John and Ogle 2009).
Seed Yields And Stand Life
Seed can be harvested in the first year following fall seeding (Shock et al. 2015). Seed yields of 75 to 400 lbs PLS/ac (84–450 kg PLS/ha) have been reported, and harvests can be made for 4 to 6 years (Stevens et al. 1996; St. John and Ogle 2009; Shock and Feibert 2016).
In seed fields growing at OSU MES, seed yield of scarlet globemallow was greatest in the fourth growing season (Shock et al. 2015). This exceeded the yield in the establishment year when multiple hand collections were made. In all but the establishment year, crops were flailed each fall to concentrate timing of flowering and reduce duration of flowering. Seed was then harvested with a single mechanical harvest with a small plot combine (Table 8; Shock et al. 2015). Plants produced harvestable seed for 5 years (Shock et al. 2015), but by the fifth-year seed production was greatly reduced. In the sixth year, growth was poor, and the stands were eliminated (Shock et al. 2015).
Table 8. Flowering dates, harvest dates, and seed yields for seed production fields provided with no supplemental irritation at Oregon State University’s Malheur Experiment Station in Ontario, OR (Shock et al. 2015).
|
Year |
Start of flowering |
End of flowering |
Harvest date(s) |
Seed yield (lbs/ac) |
|
2007 |
5 May |
Not reported |
20 June, |
279.8* |
|
2008 |
5 May |
15 June |
21 July |
298.7 |
|
2009 |
1 May |
10 June |
14 July |
332.2 |
|
2010 |
10 May |
25 June |
20 July |
385.7 |
|
2011 |
26 May |
14 July |
29 July |
89.6 |
* Seed yield represents multiple hand-harvests.
In seed production experiments conducted in Utah, seed produced per plant and per acre was greater for plants grown with fabric mulch than for those grown with plastic mulch or without mulch. Although production/plant was greater for plants spaced 12 in (30 cm) apart than those spaced 6 in (15 cm), production/ac was generally greater for plants spaced 6 in (15 cm) than those spaced 12 in (30 cm) (Anderson et al. 2006). At seed production plots at Fountain Green, Utah, almost 3 g of seed/plant was produced when plants were spaced 12 in (30 cm). A little over 4 g of seed/plant was produced when plants were spaced 18 in (46 cm) (Meyer and Vernon 2008).
Nursery Practice
Scarlet globemallow can be propagated from seed or spring-collected rhizomes (LBJWC 2018). Seedlings were grown successfully in the greenhouse when sand paper-scarified seeds were seeded in containers filled with media of four parts sand, two parts peat moss, and one part vermiculite (Pendery and Rumbaugh 1990). In a greenhouse study, spring and summer growth of scarlet globemallow was evaluated in media with a pH range of 4.5 to 8.1. Optimal growth occurred at pH levels of 5.5 to 7.2 (Cardoso et al. 2007).
Wildland Seeding And Planting
Seeding and planting methods have been used to revegetate or restore wildland sites with scarlet globemallow.

Figure 9. Scarlet globemallow seed being stored at the Utah Division of Wildlife Resources’ Great Basin Research Center until use in wildland restoration. Photo: N. Shaw, USFS.
Seeding. Fall seeding is recommended using broadcast, drill, or cultipacker seeding methods (Shaw and Monsen 1983). Seed should be covered by 0.25 in (0.6 cm) or less of soil (Wasser 1982; USDA NRCS IDPMC 2005; St. John and Ogle 2009). The pure stand seeding rate is 2 to 3 lbs PLS/ac (2.2–3.4 kg PLS/ha), but it is recommended as only about 10% of seeding mixtures (Wasser 1982; USDA NRCS IDPMC 2005; St. John and Ogle 2009). Row spacings of 2 to 4 ft (0.6–1.2 m) are recommended and seeding in alternate or selected rows should be considered if scarlet globemallow is being seeded with species with rapid establishment and early growth (Wasser 1982; Ogle et al. 2011).
In field experiments, acid-scarified seed that was drill seeded failed to establish, even though acid-scarified seed that was planted in bags germinated in the same area (James 2014, 2015). Germination within the bags was almost 70% at a wet site with sandy loam soils, a little more than 20% at a wet site with silt loam soils, and less than 10% at a mid-precipitation site with silt loam soils and a dry site with sandy loam soils. All seeding was done in the fall (James 2014, 2015).
Scarlet globemallow has been used successfully in several revegetation and restoration projects. At Scotts Bluff National Monument in Nebraska, frequency of scarlet globemallow was 8% at a golf course restoration site 1 to 2 years after being seeded at a rate of 0.24 lb/ac (0.28 kg/ha) (Huddle et al. 2001). In field experiments in Utah, scarlet globemallow survival to year 3 was greater where it was seeded in soil with intact soil crusts (27%) than where the top 0.4 in (1 cm) of soil crusts was removed (17%) (Harper cited in (Belnap 1994). On coal mine spoils at the Dickinson Experiment Station, North Dakota, there were 21 seedlings/m² (1.9/ft²) by August 1978 following a heavy rate of seeding in June 1977. Transplants were also planted at this site and it is unclear if they were included in the seedling count (Bjugstad and Whitman 1982).
Scarlet globemallow was important following seeding in disturbed Wyoming big sagebrush (Artemisia tridentata subsp. wyomingensis) at the Piceance Basin, Colorado. Experimental disturbances similar to those created by oil shale mining included removing top 2 in (5 cm) and mixing the next 10 in (25 cm) of soil and removing 90% or more of the seed bank. In the fall, disturbed sites were seeded with either a mix of early or late seral species. Seed was broadcast by hand and lightly covered with soil. Scarlet globemallow was part of the early seral seed mix and seeded at a 0.81 lb PLS/ac (0.91 kg PLS/ha) rate. Twenty five years later scarlet globemallow was most important in disturbed-unseeded and disturbed-early seral seeded plots (Table 9; (Hoelzle et al. 2012).
Table 9. Average total biomass (%) of scarlet globemallow on restoration sites in Wyoming big sagebrush vegetation in the Piceance Basin (Hoelzle et al. 2012).
|
Time since treated |
Early seral seeded* |
Late-seral seeded |
Unseeded |
Undisturbed |
|
—–average total biomass (%)—– |
||||
|
24 yrs |
1.7 |
2.9 |
3.4 |
0.5 |
|
25 yrs |
5.6 |
2.6 |
6.6 |
1.0 |
* Scarlet globemallow a part of the early seral seed mix. Seeded at 0.91 kg PLS/ha rate.
Scarlet globemallow failed to emerge in the first 2 seasons after being seeded at an arid desert shrub site at McInnis Canyon National Conservation Area, Colorado, where annual precipitation is just 3 to 5 in (8–13 cm). The site was herbicide treated to reduce competition from annual grasses a year before seeding with a cone seeder on August 24. Emergence was limited for most seeded forbs (Grant-Hoffman et al. 2015).
Planting. Establishment is generally expected to be good for bareroot/wilding and container grown stock of scarlet globemallow (Stevens 2004). Several studies report high establishment when using plants in wildland settings. Overall survival was high (86%) a year after 3-month-old scarlet globemallows were planted in outdoor dryland pastures for livestock grazing studies in southern Idaho and northern Utah (Pendery and Rumbaugh 1990). After 3 months in a greenhouse, stock from 4 scarlet globemallow seed accessions was planted into clean-tilled sites in mid-April 1987. By summer 1988, transplants averaged 33 grams (range: 14–42 g), 13 stems (range: 10-17), and had a maximum stem length average of 7 in (19 cm) (range: 5–9 in [12-23 cm]). Between late June and mid-September 1988, 15% of schizocarps were ripe. Plant biomass (for all Sphaeralcea spp. evaluated in this study) was greater, plants flowered earlier, and seed production was greater at the northern Utah than the southern Idaho site, where annual precipitation was 2 to 16 in (59–405 mm) greater between 1985 and 1988 (Pendery and Rumbaugh 1990).
Transplant survival was nearly 100% when 6-month-old plants were used on coal mine spoils at the Dickinson Experiment Station, North Dakota. Transplants were 4 in (11 cm) tall at the end of August following June planting. Flowers were produced by transplants (Bjugstad and Whitman 1982).
Longer-term assessments in salt desert (Pendleton et al. 1992) and on bentonite mine sites (Uresk and Yamamoto 1994), revealed poor persistence of scarlet globemallow beyond the fourth or fifth post-planting years. Survival was 94% in the second year, 65% in the third, 27% in the fourth, and 13% in the fifth year following planting of scarlet globemallow in a salt desert shrubland in Rush Valley, Utah. Seed collected from an unknown location was grown for 6 months before being transplanted in early May 1977. Soils at the study site included a slightly saline surface layer over fine sandy or silt loams. No scarlet globemallow plants occurred in the sixth year of monitoring (Pendleton et al. 1992). Scarlet globemallow survival was low (5%) 4 years after 1-year-old plants were planted on bentonite spoils near Upton, Wyoming. Surviving plants were only found on spoils amended with vermiculite and did not occur on unamended spoils, which were described as saline and sodic prior to any amendments (Uresk and Yamamoto 1994).
Stand management. Newly seeded or planted sites should be protected from grazing and trampling for at least 2 years (Wasser 1982; St. John and Ogle 2009). Sites may also need protection from birds and rabbits when populations are high, since they can be heavy seed predators (Pendery and Rumbaugh 1986). Once stands are established, grazing should include rest periods during the growing season to encourage persistence through self-seeding (Wasser 1982).
Studies indicate that at least short-term decreases were associated with herbicide treatments in wildland settings. Abundance and size of scarlet globemallow plants were reduced where imazapic was applied at a high rate (0.03 lb/ac [0.175 kg/ha]) to mowed Wyoming big sagebrush vegetation in southwestern Colorado. Cover of scarlet globemallow increased 1.6% on control and decreased 3.4% on treated sites 1 year after herbicide applications. Average height increased 1.3 in (3.2 cm) on control and decreased 0.6 in (1.6 cm) on treated sites (Baker et al. 2009). Frequency of scarlet globemallow was 17% lower on treated than control plots following the third late fall application of atrazine (1.75 lb/ac [2 kg/ha]) at the Central Plains Experimental Range (P = 0.05) (Houston 1977).
Acknowledgements
Funding for Western Forbs: Biology, Ecology, and Use in Restoration was provided by the USDI BLM Great Basin Native Plant Materials Ecoregional Program through the Great Basin Fire Science Exchange. Great thanks to the chapter reviewers: Lesley DeFalco, USGS, and Rosemary Pendelton, USFS-retired.
This research was supported in part by the USDA Forest Service, Rocky Mountain Research Station. The findings and conclusions in this publication are those of the authors and should not be construed to represent any official USDA or U.S. Government determination or policy.
Literature Cited
Albertson, F.W. 1937. Ecology of mixed prairie in west central Kansas. Ecological Monographs. 7(4): 483-547.
Albertson, F.W.; Tomanek, G.W.; Riegel, A. 1957. Ecology of drought cycles and grazing intensity on grasslands of central Great Plains. Ecological Monographs. 27(1): 27-44.
Albertson, F.W.; Weaver, J.E. 1944. Nature and degree of recovery of grassland from the great drought of 1933 to 1940. Ecological Monographs. 14(4): 394-479.
Alward, R.D.; Detling, J.K.; Milchunas, D.G. 1999. Grassland vegetation changes and nocturnal global warming. Science. 283(5399): 229-231.
Anderson, V.J.; Johnson, R.; Roundy, B. 2006. Agronomic and cultural care of wildland plants. In: Shaw, N.; Pellant, M., eds. Great Basin Native Plant Project: 2005 Progress Report. Boise, ID: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station: 25-28.
Armentrout, S.M.; Pieper, R.D. 1988. Plant distribution surrounding Rocky Mountain pinyon pine and oneseed juniper in south-central New Mexico. Journal of Range Management. 41(2): 139-143.
Arthun, D.; Holechek, J.L.; Wallace, J.D.; Galyean, M.L.; Cardenas, M.; Rafique, S. 1992b. Forb and shrub influences on steer nitrogen retention. Journal of Range Management. 45(2): 133-136.
Arthun, D.; Holechek, J.L.; Wallace, J.D.; Gaylean, M.L.; Cardenas, M. 1992a. Forb and shrub effects on ruminal fermentation in cattle. Journal of Range Management 45(6): 519-522.
Association of Official Seed Analysts [AOSA]. 2010. AOSA/SCST tetrazolium testing handbook. Contribution No. 29. Lincoln, NE: Association of Official Seed Analysts.
Association of Official Seed Analysts [AOSA]. 2016. AOSA rules for testing seeds. Vol. 1. Principles and procedures. Washington, DC: Association of Official Seed Analysts.
Atwood, N.D.; Welsh, S.L. 2002. Overview of Sphaeralcea (Malvaceae) in southern Utah and northern Arizona, U.S.A., and description of a new species. Novon. 12(2): 159-166.
Augustine, D.J.; Milchunas, D.G.; Derner, J.D. 2013. Spatial redistribution of nitrogen by cattle in semiarid rangeland. Rangeland Ecology and Management. 66(1): 56-62.
Ault, S.C.; Stormer, F.A. 1983. Seasonal food selection by scaled quail in northwest Texas. The Journal of Wildlife Management. 47(1): 222-228.
Baker, W.L.; Garner, J.; Lyon, P. 2009. Effect of imazapic on cheatgrass and native plants in Wyoming big sagebrush restoration for Gunnison sage-grouse. Natural Areas Journal. 29(3): 206-209.
Barner, J. 2009. Propagation protocol for production of propagules (seeds, cuttings, poles, etc.) Sphaeralcea coccinea (Nutt.) Rydb. seeds. Native Plant Network. U.S. Department of Agriculture, Forest Service, National Center for Reforestation, Nurseries, and Genetic Resources. http://npn.rngr.net/propagation/protocols [Accessed 2018 June 13].
Basey, A.C.; Fant, J.B.; Kramer, A.T. 2015. Producing native plant materials for restoration: 10 rules to collect and maintain genetic diversity. Native Plants Journal. 16(1): 37-53.
Belcher, E. 1985. Handbook on seeds of browse-shrubs and forbs. Tech. Pub. No. R8-TP8. Atlanta, GA: U.S. Department of Agriculture, Forest Service, Southern Region. 246 p.
Belnap, J. 1994. Potential role of cryptobiotic soil crusts in semiarid rangelands. In: Monsen, S.B.; Kitchen, S.G., comps. Proceedings – Ecology and management of annual rangelands; 1992 May 18-22; Boise, ID. Gen. Tech. Rep INT-GTR-313. Ogden, UT: U.S. Department of Agriculture, Forest Service, Intermountain Research Station: 179-185.
Bender, L.C.; Lomas, L.A.; Kamienski, T. 2007. Habitat effects on condition of doe mule deer in arid mixed woodland-grassland. Rangeland Ecology and Management. 60(3): 277-284.
Biondini, M.E.; Bonham, C.D.; Redente, E.F. 1985. Secondary successional patterns in a sagebrush (Artemisia tridentata) community as they relate to disturbance and soil biological activity. Vegetatio. 60(1): 25-36.
Bjugstad, A.J.; Whitman, W.C. 1982. Perennial forbs for wildlife habitat restoration on mined lands in the northern Great Plains. In: Proceedings – 62nd annual conference of the Western Association of Fish and Wildlife Agencies. 1982 July 19-22; Las Vegas, NV. Cheyenne, WY: Western Association of Fish and Wildlife Agencies: 257-271.
Bock, J.H.; Bock, C.E.; O’Shea Stone, M. 1986. The effects of fire on virgin northern mixed grassland at Custer Battlefield National Monument. University of Wyoming National Park Service Research Center Annual Report. 10(7): 43-55.
Bonham, C.D.; Lerwick, A. 1976. Vegetation changes induced by prairie dogs on shortgrass range. Journal of Range Management. 29(3): 221-225.
Bower, A.D.; St. Clair, J.B.; Erickson, V. 2014. Generalized provisional seed zones for native plants. Ecological Applications. 24(5): 913-919.
Brockway, D.G.; Gatewood, R.G.; Paris, R.B. 2002. Restoring fire as an ecological process in shortgrass prairie ecosystems: Initial effects of prescribed burning during the dormant and growing season. Journal of Environmental Management. 65(2): 135-152.
Buechner, H.K. 1950. Life history, ecology, and range use of the pronghorn antelope in Trans-Pecos Texas. American Midland Naturalist. 43(2): 257-354.
Busby, R.R.; Gebhart, D.L.; Stromberger, M.E.; Meiman, P.J.; Paschke, M.W. 2011. Early seral plant species interactions with an arbuscular mycorrhizal fungi community are highly variable. Applied Soil Ecology. 48(3): 257-262.
Cain, J.W.; Avery, M.M.; Caldwell, C.A.; Abbott, L.B.; Holechek, J.L. 2017. Diet composition, quality and overlap of sympatric American pronghorn and gemsbok. Wildlife Biology. 1: 1-10.
Callow, J.M.; Kantrud, H.A.; Higgins, K.H. 1992. First flowering dates and flowering periods of prairie plants at Woodworth, North Dakota. Prairie Naturalist. 24(2): 57-64.
Cane, J.H. 2008. Pollinating bees crucial to farming wildflower seed for U.S. habitat restoration. In: James, R.; Pitts-Singer, T., eds. Bees in agricultural ecosystems. Oxford, UK: Oxford University Press: 48-64.
Cane, J.H. 2009. Pollination and seed predator studies. In: Shaw, N.; Pellant, M., eds. Great Basin Native Plant Project: 2008 Progress Report. Boise, ID: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station: 62-65.
Cardoso, G.; Cerny-Koenig, T.; Koenig, R.; Kjelgren, R. 2007. Characterizing fertilizer and media pH requirements for greenhouse production of Intermountain West native herbaceous perennials. Native Plants Journal. 8(2): 115-121.
Chamberlin, R.V. 1911. The ethno-botany of the Gosiute Indians. American Anthropological Association. 63(1): 329-405.
Coffin, D.P.; Urban, D.L. 1993. Implications of natural history traits to system-level dynamics: Comparisons of a grassland and a forest. Ecological Modelling. 67(2-4): 147-178.
Colorado State University Extension [CSU Ext.]. 2016. Western Colorado insects: Native plant seed production. http://www.wci.colostate.edu/seed_production.shtml
Consortium of Pacific Northwest Herbaria [CPNWH]. 2017. Seattle, WA: University of Washington Herbarium, Burke Museum of Natural History and Culture. pnwherbaria.org/
Costello, D.F. 1944. Important species of the major forage types in Colorado and Wyoming. Ecological Monographs. 14(1): 107-134.
Costello, D.F. 1944a. Natural revegetation of abandoned plowed land in the mixed prairie association of northeastern Colorado. Ecology. 25(3): 312-326.
Coupland, R.T. 1950. Ecology of mixed prairie in Canada. Ecological Monographs. 20(4): 271-315.
Coupland, R.T.; Johnson, R.E. 1965. Rooting characteristics of native grassland species in Saskatchewan. Journal of Ecology. 53(2): 475-507.
Craighead, J.J.; Craighead, F.C. Jr.; Davis, R.J. 1963. A field guide to Rocky Mountain wildflowers from northern Arizona and New Mexico to British Columbia. Boston, MA: Houghton Mifflin Company. 277 p.
Daubenmire, R. 1952. Forest vegetation of northern Idaho and adjacent Washington, and its bearing on concepts of vegetation classification. Ecological Monographs. 22(4): 301-330.
deCalesta, D.S. 1979. Spring and summer foods of Audubon’s cottontail rabbit (Sylvilagus audubonii) in north-central Colorado. Southwestern Naturalist. 24(3): 549-553.
Dickinson, C.E.; Dodd, J.L. 1976. Phenological pattern in the shortgrass prairie. American Midland Naturalist. 96(2): 367-378.
Dix, R.L. 1958. Some slope-plant relationships in the grasslands of the Little Missouri Badlands of North Dakota. Journal of Range Management. 11(2): 88-92.
Dix, R.L. 1960. The effects of burning on the mulch structure and species composition of grasslands in western North Dakota. Ecology. 41(1): 49-56.
Dunn, B. 2011. Improved germination of two Sphaeralcea A. St.-Hil. (Malvaceae) species with scarification plus stratification treatments. Native Plants Journal. 12(1): 13-17.
Ellis, J.E.; Travis, M. 1975. Comparative aspects of foraging behaviour of pronghorn antelope and cattle. Journal of Applied Ecology. 12(2): 411-420.
Elmore, F.H. 1944. Ethnobotany of the Navajo: A monograph of the University of New Mexico and the School of American Research No. 8. Santa Fe, NM: University of New Mexico. 136 p.
Erichsen-Arychuk, C.; Bork, E.W.; Bailey, A.W. 2002. Northern dry mixed prairie responses to summer wildlife and drought. Journal of Range Management. 55(2): 164-170.
European Native Seed Conservation Network [ENSCONET]. 2009. ENSCONET seed collecting manual for wild species. Edition 1: 32 p.
Fagerstone, K.A.; Tietjen, H.P.; Williams, O. 1981. Seasonal variation in the diet of black-tailed prairie dogs. Journal of Mammology. 62(4): 820-824.
Fahnestock, J.T.; Detling, J.K. 2002. Bison-prairie dog-plant interactions in a North American mixed-grass prairie. Oecologia. 132(1): 86-95.
Farr, D.F.; Rossman, A.Y. 2017. Fungal databases, U.S. National Fungus Collections. U.S. Department of Agriculture, Agricultural Research Service. https://nt.ars-grin.gov/fungaldatabases/.
Fisher, J.T.; Fancher, G.A.; Aldon, E.F. 1990. Factors affecting establishment of one-seed juniper (Juniperus monosperma) on surface-mined lands in New Mexico. Canadian Journal of Forest Research 20: 880-886.
Flake, L.D. 1973. Food habits of four species of rodents on a short-grass prairie in Colorado. Journal of Mammalogy. 54(3): 636-647.
Flinders, J.T.; Crawford, J.A. 1977. Composition and degradation of jackrabbit and cottontail fecal pellets, Texas High Plains. Journal of Range Management. 30(3): 217-220.
Goetz, H. 1963. Growth and development of native range plants in the mixed grass prairie of western North Dakota. Fargo, ND: North Dakota State University. Thesis. 165 p.
Goetz, H. 1969. Composition and yields of native grassland sites fertilized at different rates of nitrogen. Journal of Range Management. 22(6): 384-390.
Gokbulak, F. 2002. Effect of American bison (Bison bison L.) on the recovery and germinability of seeds of range forage species. Grass and Forage Science. 57(4): 395-400.
Grant-Hoffman, N.; Parr, S.; Blanke, T. 2015. Native plant species field evaluation in salt desert: Good materials, bad situation. Native Plants Journal. 16(2): 87-95.
Grygiel, C.E.; Bonham, C.D.; Redente, E.F. 1984. Combined effects of environmental and agronomic factors on the invasion patterns of Sphaeralcea coccinea (Nutt.) Rydb. (Malvacea). Phytologia. 56(3): 145-153.
Halvorson, G.A.; Lang, K.J. 1989. Revegetation of a salt water blowout site. Journal of Range Management. 42(1): 61-65.
Hammon, R.; Franklin, M. 2012. Seed-feeding insects impacting globemallow seed production. Native Plants Journal. 13(2): 95-97.
Hansen, R.M.; Gold, I.K. 1977. Blacktail prairie dogs, desert cottontails and cattle trophic relations on shortgrass range. Journal of Range Management. 30(3): 210-214.
Hanson, H.C. 1955. Characteristics of the Stipa comata–Bouteloua gracilis–Bouteloua curtipendula association in northern Colorado. Ecology. 36(2): 269-280.
Hanson, H.C.; Dahl, E. 1957. Some grassland communities in the mountain-front zone in northern Colorado. Vegetatio. 7(4): 249-270.
Hart, J.A. 1981. The ethnobotany of the northern Cheyenne Indians of Montana. Journal of Ethnopharmacology. 4(1): 1-55.
Hatton, T.; West, N. 1987. Early seral trends in plant community diversity on a recontoured surface mine. Vegetatio. 73(1): 21-29.
Haufler, J.B.; Nagy, J.G. 1984. Summer food habits of a small mammal community in the pinyon-juniper ecosystem. The Great Basin Naturalist. 44(1): 145-150.
Havstad, K.; Pieper, R.D.; Donart, G.B.; Wallace, J.D.; Cordova, F.J.; Parker, E.E. 1979. Cattle diets on a fertilized blue grama upland range site. Journal of Range Management. 32(5): 398-401.
Henderson, C.B. 1990. The influence of seed apparency, nutrient content and chemical defenses on dietary preference in Dipodomys ordii. Oecologia. 82(3): 333-341.
Hermann, F. 1966. Notes on western range forbs: Cruciferae through Compositae. Agric. Handb. 293. Washington, DC: U.S. Department of Agriculture, Forest Service. 365 p.
Hewitt, G.B. 1980. Plant phenology as a guide in timing grasshopper control efforts on Montana USA rangeland. Journal of Range Management. 33(4): 297-299.
Hladek, K.L.; Hulett, G.K.; Tomanek, G.W. 1972. The vegetation of remnant shale-limestone prairies in western Kansas. The Southwestern Naturalist. 17(1): 1-10.
Hoelzle, T.B.; Jonas, J.L.; Paschke, M.W. 2012. Twenty-five years of sagebrush steppe plant community development following seed addition. Journal of Applied Ecology. 49(4): 911-918.
Holmgren, N.H.; Holmgren, P.K.; Cronquist, A. 2005. Intermountain flora: Vascular plants of the Intermountain West, U.S.A. Volume Two, Part B: Subclass Dilleniidae. Bronx, NY: New York Botanical Garden Press. 488 p.
Houston, W.R. 1977. Species susceptibility to atrazine herbicide on shortgrass range. Journal of Range Management. 30(1): 50-52.
Houston, W.R.; Hyder, D.N. 1975. Ecological effects and fate of N following massive N fertilization of mixed-grass plains. Journal of Range Management. 28(1): 56-60.
Howe, G.; St, Clair, B.; Bachelet, D. 2017. Seedlot Selection Tool. Corvallis, OR: Conservation Biology Institute. https://seedlotselectiontool.org/sst/
Huddle, J.A.; Tichota, G.R.; Stubbendieck, J.; Stumpf, J.A. 2001. Evaluation of grassland restoration at Scotts Bluff National Monument. In: Bernstein, N.P.; Ostrander, L.J., eds. Seeds for the future; Roots of the past: Proceedings of the 17th North American Prairie Conference; 2000 July 16-20. Mason City, IA: North Iowa Community College: 125-135.
Hulett, G.K.; Sloan, C.D.; Tomanek, G.W. 1968. The vegetation of remnant grasslands in the loessial region of northwestern Kansas and southwestern Nebraska. The Southwestern Naturalist. 13(4): 377-391.
Hyder, D.N.; Bement, R.E.; Remmenga, E.E.; Terwilliger, C. 1966. Vegetation-soils and vegetation-grazing relations from frequency data. Journal of Range Management. 19(1): 11-17.
James, J. 2014. The ecology of Great Basin native forb establishment. In: Kilkenny, F.; Shaw, N.; Gucker, C., eds. Great Basin Native Plant Project: 2013 Progress Report. Boise, ID: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station: 183-187.
James, J. 2015. The ecology of Great Basin native forb establishment. In: Kilkenny, F.; Halford, A.; Malcomb, A., eds. Great Basin Native Plant Project: 2014 Progress Report. Boise, ID: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station: 168-174.
Johnson, J.R.; Nichols, J.T. 1970. Plants of South Dakota grasslands: A photographic study. Brookings, SD: South Dakota State University, Agricultural Experiment Station. 163 p.
Johnson-Nistler, C.M.; Sowell, B.F.; Sherwood, H.W.; Wambolt, C.L. 2004. Black-tailed prairie dog effects on Montana’s mixed-grass prairie. Journal of Range Management. 57(6): 641-648.
Johnston, A. 1970. Blackfoot Indian utilization of the flora of the northwestern Great Plains. Economic Botany. 24(3): 301-324.
Jones, C.; Whittaker, A. 2011. Native plant material development and cultivation trials for native Great Basin forbs and grasses. In: Shaw, N.; Pellant, M., eds. Great Basin Native Plant Selection and Increase Project 2010 Progress Report. Boise, ID: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station: 57-63.
Jorgensen, K.R.; Stevens, R. 2004. Seed collection, cleaning, and storage. In: Monsen, S.B.; Stevens, R.; Shaw, N.L., comps. Restoring western ranges and wildlands, vol. 3. Gen. Tech. Rep. RMRS-GTR-136-vol-3. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station: 699-716.
Kemp, P.R. 1983. Phenological patterns of Chihuahuan Desert plants in relation to the timing of water availability. Journal of Ecology. 71(2): 427-436.
Kildisheva, O.A.; Dumroese, R.K.; Davis, A.S. 2013. Boiled, tumbled, burned, and heated: Seed scarification techniques for Munro’s globemallow appropriate for large-scale application. Native Plants Journal. 14(1): 43-47.
Kirby, D.R.; Pessin, M.F.; Clambey, G.K. 1986. Disappearance of forage under short duration and seasonlong grazing. Journal of Range Management. 39(6): 496-500.
Kitchen, S.G. 1994. Perennial forb life-history strategies on semiarid rangelands: Implications for revegetation. In: Monsen, S.B.; Kitchen, S.G., comps. Proceedings – Ecology and management of annual rangelands; 1992 May 18-22; Boise, ID. Gen. Tech. Rep INT-GTR-313. Ogden, UT: U.S. Department of Agriculture, Forest Service, Intermountain Research Station: 342-346.
Koerth, B.H.; Krysl, L.J.; Sowell, B.F.; Bryant, F.C. 1984. Estimating seasonal diet quality of pronghorn antelope from fecal analysis. Journal of Range Management. 37(6): 560-564.
Kramer, A.T.; Foxx, A. 2016. Propagation protocol for production of propagules (seeds, cuttings, poles, etc.) Sphaeralcea coccinea seeds. Native Plant Network. U.S. Department of Agriculture, Forest Service, National Center for Reforestation, Nurseries, and Genetic Resources. http://npn.rngr.net/propagation/protocols [Accessed 2018 June 13].
La Duke, J. 2016. 48. Sphaeralcea. In: Flora of North America Editorial Committee, ed. Flora of North America North of Mexico. Volume 6 Malvaceae. http://www.efloras.org
La Duke, J.C.; Northington, D.K. 1978. The systematics of Sphaeralcea coccinea (Nutt.) Rydb. (Malvaceae). Southwestern Naturalist. 23(4): 651-660.
Lady Bird Johnson Wildflower Center [LBJWC]. 2018. Sphaeralcea coccinea (Nutt.) Rydb. Native Plant Database. Austin, TX: Lady Bird Johnson Wildflower Center. https://www.wildflower.org/plants-main [Accessed 2018 August 2].
Lauenroth, W.K.; Adler, P.B. 2008. Demography of perennial grassland plants: Survival, life expectancy and life span. Journal of Ecology. 96(5): 1023-1032.
Ley, E.; Buchmann, S.; Stritch, L.; Soltz, G. 2007. Selecting plants for pollinators: A regional guide for farmers, land managers, and gardeners in the Intermountain Semidesert Province. San Francisco, CA: Pollinator Partnership. 24 p.
Looman, J. 1963. Preliminary classification of grasslands in Saskatchewan. Ecology. 44(1): 15-29.
Magle, S.B.; Crooks, K.R. 2008. Interactions between black-tailed prairie dogs (Cynomys ludovicianus) and vegetation in habitat fragmented by urbanization. Journal of Arid Environments. 72(3): 238-246.
Mahgoub, E.E. 1984. Seasonal food habits of mule deer in the foothills of the Sacramento Mountains, New Mexico. Las Cruces, NM: New Mexico State University. Thesis. 89 p.
Marquiss, R.; Lang, R. 1959. Vegetational composition and ground cover of two natural relict areas and their associated grazed areas in the Red Desert of Wyoming. Journal of Range Management. 12(3): 104-109.
McArthur, E.D.; Young, S.A. 1999. Development of native seed supplies to support restoration of pinyon-juniper sites. In: Monsen, S.B.; Stevens, R., eds. Ecology and management of pinyon-juniper communities within the Interior West: Sustaining and restoring a diverse ecosystem. Provo, UT: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station: 327-330.
McLendon, T.; Redente, E.F. 1991. Nitrogen and phosphorus effects on secondary succession dynamics on a semi-arid sagebrush site. Ecology. 72(6): 2016-2024.
McLendon, T.; Redente, E.F. 1992. Effects of nitrogen limitation on species replacement dynamics during early secondary succession on a semiarid sagebrush site. Oecologia. 91(3): 312-317.
Mee, W.; Barnes, J.; Kjelgren, R.; Sutton, R.; Cerny, T.; Johnson, C. 2003. Water wise: Native plants for Intermountain landscapes. Logan, UT: Utah State University Press. 220 p.
Meier, G.; Weaver, T. 1997. Desirables and weeds for roadside management – A northern Rocky Mountain catalogue. Report No. RHWA/MT-97/8115. Final report: July 1994-December 1997. Helena, MT: State of Montana Department of Transportation, Research, Development, and Technology Transfer Program. 145 p.
Menke, J.W.; Trlica, M.J. 1981. Carbohydrate reserve, phenology, and growth cycles of nine Colorado range species. Journal of Range Management. 34(4): 269-277.
Meyer, T.; Vernon, J. 2008. Native plant material development and seed and seeding technology for native Great Basin forbs and grasses. In: Shaw, N.; Pellant, M., eds. Great Basin Native Plant Selection and Increase Project: 2007 Progress Report. Boise, ID: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station: 7-24.
Milchunas, D.G.; Lauenroth, W.K.; Chapman, P.L.; Kazempour, M.K. 1989. Effects of grazing, topography, and precipitation on the structure of a semiarid grassland. Vegetatio. 80(1): 11-23.
Milchunas, D.G.; Noy-Meir, I. 2004. Geologic grazing refuges and grassland diversity: A shortgrass steppe study. Journal of Range Management. 57(2): 141-147.
Moerman, D. 2003. Native American ethnobotany: A database of foods, drugs, dyes, and fibers of Native American peoples, derived from plants. Dearborn, MI: University of Michigan. http://naeb.brit.org
Mueggler, W.F.; Stewart, W.L. 1980. Grassland and shrubland habitat types of western Montana. Gen. Tech. Rep. INT-66. Ogden, UT: U.S. Department of Agriculture, Forest Service, Intermountain Forest and Range Experiment Station. 154 p.
Munson, S.M.; Lauenroth, W.K. 2009. Plant population and community responses to removal of dominant species in the shortgrass steppe. Journal of Vegetation Science. 20(2): 224-232.
Myers, G.T.; Vaughan, T.A. 1964. Food habits of the plains pocket gopher in eastern Colorado. Journal of Mammalogy. 45(4): 588-598.
Nelson, G.H.; Westcott, R.L. 1995. Three new species of Acmaeodera Eschscholtz (Coleoptera: Buprestidae) from the United States and Mexico. The Coleopterists Bulletin. 49(1): 77-87.
Nicholson, R.A.; Hulett, G.K. 1969. Remnant grassland vegetation in the central Great Plains of North America. Journal of Ecology. 57(3): 599-612.
Ogle, D.; Tilley, D.; Cane, J.; St. John, L.; Fullen, K.; Stannard, M.; Pavek, P. 2011. Plants for pollinators in the Intermountain West. Plant Materials Tech. Note 2A. Boise, ID: U.S. Department of Agriculture, Natural Resources Conservation Service. 40 p.
Omernik, J.M. 1987. Ecoregions of the conterminous United States. Map (scale 1:7,500,000). Annals of the Association of American Geographers. 77(1): 118-125.
Pavek, P.; Erhardt, B.; Heekin, T.; Old, R. 2012. Forb seedling identification guide for the Inland Northwest: Native, introduced, invasive, and noxious species. Pullman, WA: U.S. Department of Agriculture, Natural Resources Conservation Service, Pullman Plant Materials Center. 144 p.
Peden, D.G. 1976. Botanical composition of bison diets on shortgrass plains. The American Midland Naturalist. 96(1): 225-229.
Pendery, B.M.; Rumbaugh, M.D. 1986. Globemallows: Forbs for Utah rangelands. Utah Science: 41-45.
Pendery, B.M.; Rumbaugh, M.D. 1990. Survival and growth of globemallow (Sphaeralcea) species in dry land spaced-plant nurseries. Journal of Range Management. 43(5): 428-432.
Pendery, B.M.; Rumbaugh, M.D. 1993. Globemallows. Rangelands. 15(3): 127-130.
Pendleton, R.L.; Frischknecht, N.C.; McArthur, E.D. 1992. Long-term survival of 20 selected plant accessions in a Rush Valley, Utah, planting. Res. Note INT-403. Odgen, UT: U.S. Department of Agriculture, Forest Service, Intermountain Research Station. 7 p.
Pessin, M.F.; Kirby, D.R.; Clambey, G.K. 1986. Forage preferences exhibited by cattle on short duration grazing system. Farm Research. 43(6): 12-15.
Pitts, J.S.; Bryant, F.C. 1987. Steer and vegetation response to short duration and continuous grazing. Journal of Range Management. 40(5): 386-389.
Plant Conservation Alliance [PCA]. 2015. National seed strategy for rehabilitation and restoration 2015-2020. Washington, DC: U.S. Department of the Interior, Bureau of Land Management. 52 p.
Rawlins, J.K.; Anderson, V.J.; Johnson, R.; Krebs, T. 2009. Optimal seeding depth of five forb species from the Great Basin. Native Plants Journal. 10(1): 33-42.
Rice, R.W.; Cundy, D.R.; Weyerts, P.R. 1971. Botanical and chemical composition of esophageal and rumen fistula samples of sheep. Journal of Range Management. 24(2): 121-124.
Roth, T.E.; Holechek, J.L.; Hussain, M.Y. 1987. Germination response of three globemallow species to chemical treatment. Journal of Range Management. 40(2): 173-175.
Royal Botanic Gardens Kew [RBG Kew]. 2017. Seed Information Database (SID). Version 7.1. http://data.kew.org/sid/
Rumbaugh, M.D.; Mayland, H.F.; Pendery, B.M.; Shewmaker, G.E. 1993a. Element concentrations in globemallow herbage. Journal of Range Management. 46(2): 114-117.
Rumbaugh, M.D.; Mayland, H.F.; Pendery, B.M.; Shewmaker, G.E. 1993b. Utilization of globemallow (Sphaeralcea) taxa by sheep. Journal of Range Management 46(2): 103-109.
Rumbaugh, M.D.; Pendery, B.M.; Mayland, H.F.; Shewmaker, G.E. 1993c. Registration of ARS-2936 scarlet globemallow germplasm. Crop Science. 33(5): 1106-1108.
Runyon, N.R. 1947. The chemical composition of forbs in the native pastures at Hays, Kansas. Transactions of the Kansas Academy of Science. 49(4): 441-443.
Sampangi, R.K.; Mohan, S.K.; Shock, C.C. 2012. Etiology, epidemiology and management of diseases of native wildflower seed production. In: Shaw, N.L.; Pellant, M., eds. Great Basin Native Plant Project: 2011 Progress Report. Boise, ID: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station: 133-134.
Samuel, M.J.; Hart, R.H. 1994. Sixty-one years of secondary succession on rangelands of the Wyoming high plains. Journal of Range Management. 47(3): 184-191.
Samuel, M.J.; Hart, R.H. 1998. Nitrogen fertilization, botanical composition and biomass production on mixed-grass rangeland. Journal of Range Management. 51(4): 408-416.
Sandoval, L.; Holechek, J.; Biggs, J.; Valdez, R.; VanLeeuwen, D. 2005. Elk and mule deer diets in north-central New Mexico. Rangeland Ecology and Management. 58(4): 366-372.
Schade, J.D.; Sponseller, R.; Collins, S.L.; Stiles, A. 2003. The influence of Prosopis canopies on understorey vegetation: Effects of landscape position. Journal of Vegetation Science. 14(5): 743-750.
Schwartz, C.C.; Nagy, J.G. 1976. Pronghorn diets relative to forage availability in northeastern Colorado. The Journal of Wildlife Management. 40(3): 469-478.
SEINet – Regional Networks of North American Herbaria Steering Committee [SEINet]. 2017. SEINet Regional Networks of North American Herbaria. https://symbiota.org/seinet
Sexson, M.L.; Choate, J.R.; Nicholson, R.A. 1981. Diet of pronghorn in western Kansas. Journal of Range Management. 34(6): 489-493.
Shaw, N.; Monsen, S.B. 1983. Nonleguminous forbs for rangeland sites. In: Monsen, S.B; Shaw, N. Managing Intermountain rangelands-Improvement of range and wildlife habitats; 1981 September 15-17; Twin Falls, ID; 1982 June 22-24; Elko, NV. Gen. Tech. Rep. INT-157. Ogden, UT: U.S. Department of Agriculture, Forest Service, Intermountain Forest and Range Experiment Station: 123-131.
Shiflet, T.N. 1994. Rangeland cover types of the United States. Denver, CO: Society for Range Management. 152 p.
Shock, C.; Feibert, E. 2016. Irrigation water requirements for native wildflower seed production. In: Kilkenny, F.; Edwards, F.; Malcomb, A., eds. Great Basin Native Plant Project: 2015 Progress Report. Boise, ID: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station: 175-176.
Shock, C.; Feibert, E.; Parris, C.; Saunders, L. 2012. Direct surface seeding strategies for establishment of Intermountain West native plants for seed production. In: Shock, C.C., ed. Malheur Experiment Station Annual Report 2011. OSU AES Ext/CrS141. Corvallis, OR: Oregon State University: 130-135.
Shock, C.C.; Feibert, E.B.G.; Rivera, A.; Saunders, L.D.; Shaw, N.; Kilkenny, F. 2015. Globemallow seed production is unresponsive to irrigation in a semi-arid environment. In: Shock, C.C., ed. Malheur Experiment Station Annual Report 2014. OSU AES Ext/ CrS152. Corvallis, OR: Oregon State University: 247-251.
Shoop, M.C.; Clark, R.C.; Laycock, W.A.; Hansen, R.M. 1985. Cattle diets on shortgrass ranges with different amounts of fourwing saltbush. Journal of Range Management. 38(5): 443-449.
Sieg, C.H.; Uresk, D.W.; Hansen, R.M. 1983. Plant-soil relationships on bentonite mine spoils and sagebrush-grassland in the northern High Plains. Journal of Range Management. 36(3): 289-294.
Sims, P.L.; Singh, J.S.; Lauenroth, W.K. 1978. The structure and function of ten western North American grasslands: I. Abiotic and vegetational characteristics. Journal of Ecology. 66(1): 251-285.
Spellenberg, R. 2001. National Audubon Society field guide to North American wildflowers: Western region, revised edition. New York, NY: Alfred A. Knopf, Inc. 862 p.
Sriladda, C.; Kratsch, H.A.; Larson, S.R.; Kjelgren, R.K. 2012. Morphological and genetic variation among four high desert Sphaeralcea species. Hortscience. 47(6): 715-720.
St. John, L.; Ogle, D. 2009. Plant guide: Scarlet globemallow (Sphaeralcea coccinea) (Nutt.) Rydb. Aberdeen, ID: U.S. Department of Agriculture, Natural Resources Conservation Service, Aberdeen Plant Materials Center. 3 p.
Stanton, N.L.; Morrison, D.; Laycock, W.A. 1984. The effect of phytophagous nematode grazing on blue grama die-off. Journal of Range Management. 37(5): 447-450.
Stephenson, T.E.; Holecheck, J.L.; Kuykendall, C.B. 1985. Drought effect on pronghorn and other ungulate diets. The Journal of Wildlife Management. 49(1): 146-151.
Stevens, O.A. 1957. Weights of seeds and numbers per plant. Weeds. 5(1): 46-55.
Stevens, R. 2004. Establishing plants by transplanting and interseeding. In: Monsen, S.B.; Stevens, R.; Shaw, N.L., comps. Restoring western ranges and wildlands, vol. 3. Gen. Tech. Rep. RMRS-GTR-136-vol-3. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station: 739-744.
Stevens, R.; Jorgensen, K.R.; Young, S.A.; Monsen, S.B. 1996. Forb and shrub seed production guide for Utah. 1996. AG 501. Logan, UT: Utah State University Extension. 52 p.
Stevens, R.; Monsen, S.B. 2004. Forbs for seeding range and wildlife habitats. In: Monsen, S.B.; Stevens, R.; Shaw, N.L., comps. Restoring western ranges and wildlands, vol. 2. Gen. Tech. Rep. RMRS-GTR-136-vol-2. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station: 425-466.
Stubbendieck, J.; Hatch, S.L.; Hirsch, K.J. 1986. North American range plants. 3rd ed. Lincoln, NE: University of Nebraska Press. 465 p.
Summers, C.A.; Linder, R.L. 1978. Food habits of the black-tailed prairie dog in western South Dakota. Journal of Range Management. 31(2): 134-136.
Swank, G.R. 1932. The ethnobotany of the Acoma and Laguna Indians. Albuquerque, NM: University of New Mexico. Thesis. 86 p.
Thetford, F.O.; Pieper, R.D.; Nelson, A.B. 1971. Botanical and chemical composition of cattle and sheep diets on pinyon-juniper grassland range. Journal of Range Management. 24(6): 425-431.
Tilley, D.; Ogle, D.; Cornforth, B. 2011. The pop test: A quick aid to estimate seed quality. Native Plants Journal. 12(3): 227-232.
Tomanek, G.W.; Albertson, F.W. 1957. Variations in cover, composition, production, and roots of vegetation on two prairies in western Kansas. Ecological Monographs. 27(3): 267-281.
Trlica, M.J.; Buwai, M.; Menke, J.W. 1977. Effects of rest following defoliations on the recovery of several range species. Journal of Range Management. 30(1): 21-27.
Turner, G.T. 1971. Soil and grazing influences on a salt-desert shrub range in western Colorado. Journal of Range Management. 24(1): 31-37.
Uresk, D.W. 1984. Black-tailed prairie dog food habits and forage relationships in western South Dakota. Journal of Range Management. 37(4): 325-329.
Uresk, D.W. 1990. Using multivariate techniques to quantitatively estimate ecological stages in a mixed grass prairie. Journal of Range Management. 43(4): 282-285.
Uresk, D.W.; Yamamoto, T. 1994. Field-study of plant-survival as affected by amendments to bentonite spoil. Great Basin Naturalist. 54(2): 156-161.
USDA Forest Service, Bend Seed Extractory [USDA FS BSE]. 2017. Nursery Management Information System Version 4.1.11. Local Source Report 34-Source Received. Bend, OR: U.S. Department of Agriculture, Forest Service, Bend Seed Extractory.
USDA Forest Service, Great Basin Native Plant Project [USFS GBNPP]. 2014. Seed weight table calculations made in-house. Report on file. Boise, ID: U.S. Department of Agriculture, Forest Service, Boise Aquatic Sciences Laboratory. Available: https://www.fs.fed.us/rm/boise/research/shrub/Links/Seedweights.pdf
USDA Forest Service, Western Wildland Environmental Threat Assessment Center [USFS WWETAC]. 2017. TRM Seed Zone Applications. Prineville, OR: U.S. Department of Agriculture, Forest Service, Western Wildland Environmental Threat Assessment Center. https://research.fs.usda.gov/pnw/products/dataandtools/seed-zone-webmap
USDA Natural Resources Conservation Service [USDA NRCS]. 2017. The PLANTS Database. Greensboro, NC: U.S. Department of Agriculture, Natural Resources Conservation Service, National Plant Data Team. https://plants.sc.egov.usda.gov/
USDA, Natural Resources Conservation Service, Aberdeen, Idaho, Plant Materials Center [USDA NRCS IDPMC] 2005. Appendix 2. In: Shaw, N.; Pellant, M., eds. Great Basin Native Plant Project: 2004 Progress Report. Boise, ID: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station: 63-75.
USDI Bureau of Land Management, Seeds of Success [USDI BLM SOS]. 2016. Bureau of Land Management technical protocol for the collection, study, and conservation of seeds from native plant species for Seeds of Success. Washington, DC: U.S. Department of the Interior, Bureau of Land Management, Plant Conservation and Restoration Program. 37 p.
USDI Bureau of Land Management, Seeds of Success [USDI BLM SOS]. 2017. Seeds of Success collection data. Washington, DC: U.S. Department of the Interior, Bureau of Land Management, Plant Conservation and Restoration Program.
USDI Environmental Protection Agency [USDI EPA]. 2017. Ecoregions. Washington, DC: U.S. Department of the Interior, Environmental Protection Agency. https://www.epa.gov/eco-research/ecoregions
USDI Geological Survey [USDI USGS]. 2017. Biodiversity Information Serving Our Nation (BISON). U.S. Geological Survey. https://www.gbif.us/
Utah Crop Improvement Association [UCIA]. 2015. How to be a seed connoisseur. Logan, UT: UCIA, Utah Department of Agriculture and Food, Utah State University and Utah State Seed Laboratory. 16 p.
Utah State University Extension [USU Ext]. 2017. Range plants of Utah: Scarlet globemallow. Logan, UT: Utah State University. https://extension.usu.edu/rangeplants/forbsherbaceous/scarlet-globemallow [Accessed 2018 June 13].
Vaughan, T.A. 1967. Food habits of the northern pocket gopher on shortgrass prairie. The American Midland Naturalist. 77(1): 176-189.
Vavra, M.; Rice, R.W.; Hansen, R.M. 1978. A comparison of esophageal fistula and fecal material to determine steer diets. Journal of Range Management. 31(1): 11-13.
Vavra, M.; Rice, R.W.; Hansen, R.M.; Sims, P.L. 1977. Food habits of cattle on shortgrass range in northeastern Colorado. Journal of Range Management. 30(4): 261-263.
Veech, J.A. 2000. Predator-mediated interactions among the seeds of desert plants. Oecologia. 124(3): 402-407.
Wagner, W.L.; Martin, W.C.; Aldon, E.F. 1978. Natural succession on strip-mined lands in northwestern New Mexico. Reclamation Review. 1: 67-73.
Wasser, C.H. 1982. Ecology and culture of selected species useful in revegetating disturbed lands in the West. FWS/OBS-82/56. Washington, D.C.: U.S. Department of the Interior, Fish and Wildlife Service, Office of Biological Services, Western Energy and Land Use Team. 347 p.
Weaver, J.E. 1958. Classification of root systems of forbs of grassland and a consideration of their significance. Ecology. 39(3): 394-401.
Weaver, J.E.; Albertson, F.W. 1943. Resurvey of grasses, forbs, and undergrowth plant parts at the end of the great drought. Ecological Monographs. 13(1): 64-117.
Weaver, T.; Gustafson, D.; Lichthardt, J. 1993. Plants colonizing disturbed areas in fifteen Rocky Mountain environments: Weeds and reclamation candidates. University of Wyoming National Park Service Research Center annual report. 17(4): 1-9.
Welsh, S.L.; Atwood, N.D.; Goodrich, S.; Higgins, L.C., eds. 1987. A Utah flora. The Great Basin Naturalist Memoir 9. Provo, UT: Brigham Young University. 894 p.
Wheeler, A.G. 2011. Arhyssus confusus Chopra and A. parvicornis (Signoret) (Hemiptera: Rhopalidae): apparent specialists on globemallow (Sphaeralcea spp.; Malvaceae), with notes on their seasonality. Proceedings of the Entomological Society of Washington. 113(2): 195-202.
White, L.M. 1995. Predicting flowering of 130 plants at 8 locations with temperature and daylength. Journal of Range Management. 48(2): 108-14.
Whitman, W.C.; Hanson, H.T.; Loder, G. 1943. Natural revegetation of abandoned fields in western North Dakota. Bull. 321. Fargo, ND: North Dakota Agricultural College, Agriculture Experiment Station. 18 p.
Winter, S.L.; Cully, J.F.; Pontius, J.S. 2002. Vegetation of prairie dog colonies and non-colonized shortgrass prairie. Journal of Range Management. 55(5): 502-508.
Wyman, L.C.; Harris, S.K. 1951. The ethnobotany of the Kayenta Navaho: An analysis of the John and Louisa Wetherill ethnobotanical collection. Publications in Biology. No. 5 Albuquerque, NM: University of New Mexico Press. 66 p.
Young, S.A.; Schrumpf, B.; Amberson, E. 2003. The Association of Official Seed Certifying Agencies (AOSCA) native plant connection. Moline, IL: AOSCA. 9 p. Available: https://seedcert.oregonstate.edu/sites/seedcert.oregonstate.edu/files/pdfs/aoscanativeplantbrochure.pdf
How to Cite
Gucker, Corey L.; Shaw, Nancy L. 2019. Scarlet globemallow (Sphaeralcea coccinea). In: Gucker, C.L.; Shaw, N.L., eds. Western forbs: Biology, ecology, and use in restoration. Reno, NV: Great Basin Fire Science Exchange. Online: https://westernforbs.org/species/scarlet-globemallow-sphaeralcea-coccinea/